
Identification, Culture Characteristics and Whole-Genome Analysis of Pestalotiopsis neglecta Causing Black Spot Blight of Pinus sylvestris var. mongolica


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of P. neglecta
2.2. Culture Characteristics of P. neglecta
2.2.1. Screening of Optimal Medium
2.2.2. Screening of Carbon and Nitrogen Sources
2.2.3. Temperature and pH Tests
2.3. Whole-Genome Sequencing and Assembly
2.3.1. DNA Extraction
2.3.2. Genome Sequencing and Assembly
2.3.3. Gene Prediction and Functional Annotation
2.4. Data Availability
3. Results
3.1. Isolation and Identification of P. neglecta
3.2. Culture Characteristics of P. neglecta YJ-3
3.3. Genome Sequencing and Assembly
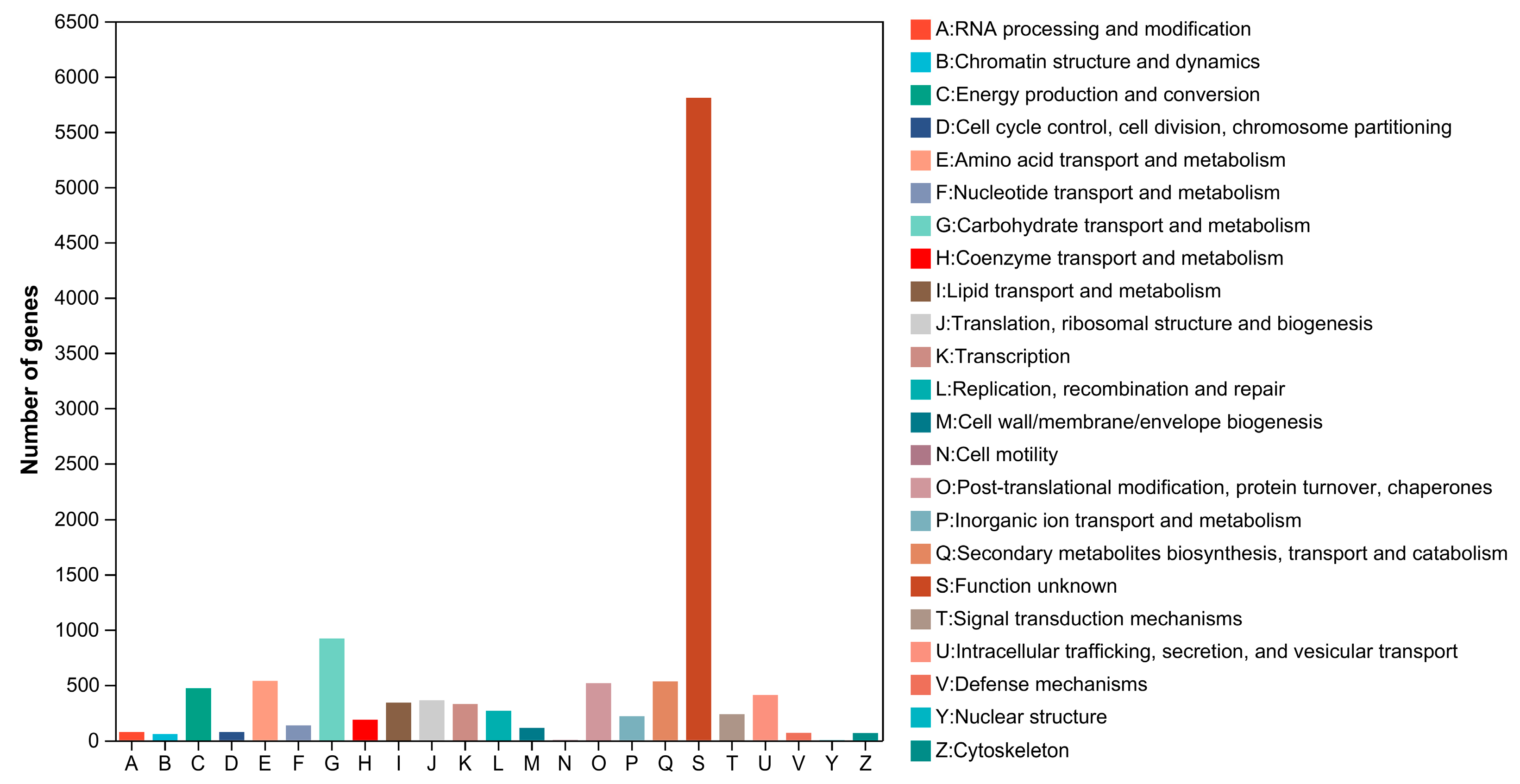
3.4. Gene Prediction and Functional Annotation
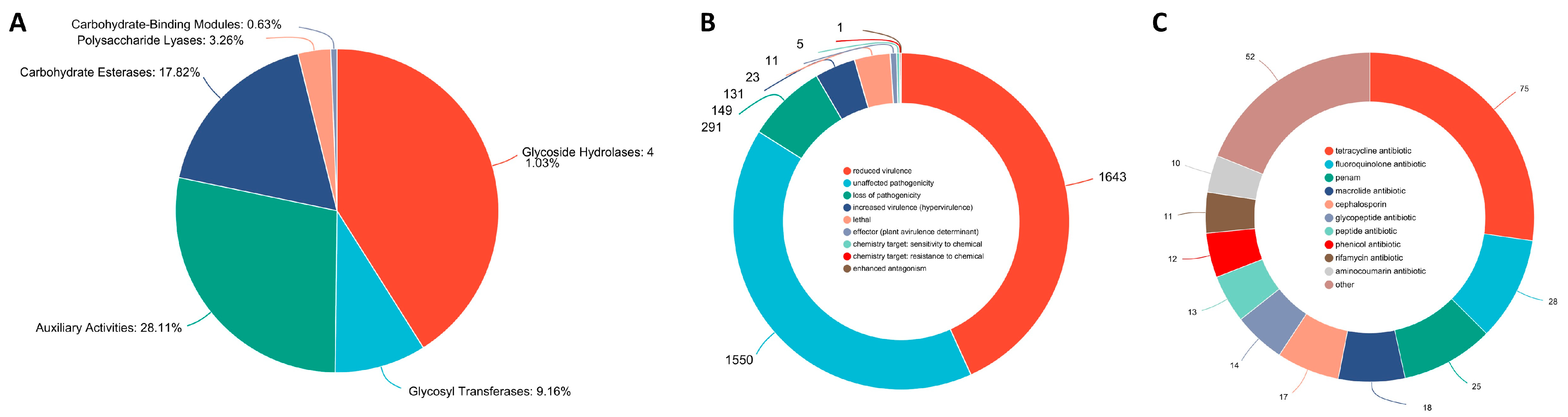
3.5. Carbohydrate-Active Enzymes (CAZymes)
3.6. Pathogenic System Analysis
3.7. Cytochromes P450
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halifu, S.; Deng, X.; Song, X.S.; An, Y.N.; Song, R.Q. Effects of Sphaeropsis Blight on Rhizosphere Soil Bacterial Community Structure and Soil Physicochemical Properties of Pinus sylvestris var. mongolica in Zhanggutai, China. Forests 2019, 10, 954. [Google Scholar] [CrossRef]
- Wang, S.-R.; Zhang, H.-X.; Chen, Y.-Z.; Zhang, Y.-D.; Li, D.-B.; Huang, Y.; Zhang, G.-C.; Yang, J. First Report of Black Spot Needle Blight of Pinus sylvestris var. mongolica Caused by Heterotruncatella spartii in China. Plant Dis. 2022, 106, 2256. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hao, X.; Liu, X.; Ma, L. First Report of Pestalotiopsis neglecta Causing Black Spot Needle Blight of Pinus sylvestris var. mongolica Litv. in China. Plant Dis. 2020, 104, 1545. [Google Scholar] [CrossRef]
- Barr, M.E. Pestalosphaeria, a new genus in the Amphisphaeriaceae. Mycologia 1975, 67, 187–194. [Google Scholar] [CrossRef]
- Barr, M.E. Prodromus to nonlichenized, pyrenomycetous members of class Hymenoascomycetes. Mycotaxon 1990, 39, 43–184. [Google Scholar]
- Kang, J.C.; Kong, R.Y.C.; Hyde, K.D. Studies on the Amphisphaeriales I. Amphisphaeriaceae (sensu stricto) and its phylogenetic relationships inferred from 5.8S rDNA and ITS2 sequences. Fungal Divers. 1998, 1, 147–157. [Google Scholar]
- Kang, J.C.; Hyde, K.D.; Kong, R.Y.C. Studies on the Amphisphaeriales. The Amphisphaeriaceae (sensu stricto). Mycol. Res. 1999, 103, 53–64. [Google Scholar] [CrossRef]
- Espinoza, J.G.; Briceno, E.X.; Keith, L.M.; Latorre, B.A. Canker and twig dieback of blueberry caused by Pestalotiopsis spp. and a Truncatella sp. in Chile. Plant Dis. 2008, 92, 1407–1414. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Yu, J.D.; Ma, C.L.; Kong, L.; He, C.Z.; Li, J. Genomic Analysis of the Mycoparasite Pestalotiopsis sp. PG52. Pol. J. Microbiol. 2021, 70, 189–199. [Google Scholar] [CrossRef]
- Zheng, S.Q.; Chen, R.Q.; Wang, Z.; Liu, J.; Cai, Y.; Peng, M.H.; Zhang, T.; Li, Y.X.; Wang, B.H.; Bao, J.D.; et al. High-Quality Genome Assembly of Pseudopestalotiopsis theae, the Pathogenic Fungus Causing Tea Gray Blight. Plant Dis. 2021, 105, 3723–3726. [Google Scholar] [CrossRef]
- Hyde, K.D.; Fröhlich, J. Mycosphaerella palmicola associated with leaf spots of Cocos nucifera in Australia Iran Jaya and Papua New Guinea. Mycol. Res. 1995, 99, 704–706. [Google Scholar] [CrossRef]
- Xu, L.; Kusakari, S.; Hosomi, A.; Toyoda, H.; Ouchi, A. Postharvest disease of grape caused by Pestalotiopsis species. Annu. Phytopathol. Soc. Jpn. 1999, 65, 305–311. [Google Scholar] [CrossRef]
- Das, R.; Chutia, M.; Das, K.; Jha, D.K. Factors affecting sporulation of Pestalotiopsis disseminata causing grey blight disease of Persea bombycina Kost., the primary food plant of muga silkworm. Crop Prot. 2010, 29, 963–968. [Google Scholar] [CrossRef]
- Ullasa, B.A.; Rawal, R.D. Occurrence of a new post-harvest disease of mango due to Pestalotiopsis glandicola. Acta Hortic. ISHS 1989, 231, 540–543. [Google Scholar] [CrossRef]
- Korsten, L.; De Jager, E.S.; De Villers, E.E.; Lourens, A.; Kotze, J.M.; Wehner, F.C. Evaluation of bacterial epiphytes isolated from avocado leaf and fruit surfaces for biocontrol of avocado postharvest diseases. Plant Dis. 1995, 79, 1149. [Google Scholar] [CrossRef]
- Tagne, A.; Mathur, S.B. First report of chlorotic spot of maize caused by Pestalotiopsis neglecta. Plant Pathol. 2001, 50, 791. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; Chen, F.; Chen, J.; Ye, J. First Report of Shoot Blight on Cryptomeria japonica Caused by Pestalotiopsis neglecta in China. Plant Dis. 2019, 103, 2140–2141. [Google Scholar] [CrossRef]
- Wang, X.N.; Zhang, X.L.; Liu, L.; Xiang, M.C.; Wang, W.Z.; Sun, X.; Che, Y.S.; Guo, L.D.; Liu, G.; Guo, L.Y.; et al. Genomic and transcriptomic analysis of the endophytic fungus Pestalotiopsis fici reveals its lifestyle and high potential for synthesis of natural products. BMC Genom. 2015, 16, 28. [Google Scholar] [CrossRef]
- Fu, F.F.; Hao, Z.D.; Wang, P.K.; Lu, Y.; Xue, L.J.; Wei, G.Y.; Tian, Y.L.; Hu, B.S.; Xu, H.B.; Shi, J.S.; et al. Genome Sequence and Comparative Analysis of Colletotrichum gloeosporioides Isolated from Liriodendron Leaves. Phytopathology 2020, 110, 1260–1269. [Google Scholar] [CrossRef]
- Czapek, F. Umtersuchung uber die sticstoffgewinnungund einweissbildung der Pfianze. Beitr. Chem. Physiol. Pathol. 1903, 1, 540. [Google Scholar]
- Dox, A.W. The Intercellular Enzymes of Penicillium and Aspergillus with Special References to Those of P. camemberti; U.S. Department of Agriculture, Bureau of Animal Industry: Atlanta, GA, USA, 1910; 70p. [Google Scholar]
- Acharya, T.; Hare, J. Sabouraud Agar and Other Fungal Growth Media. In Laboratory Protocols in Fungal Biology; Gupta, V.K., Tuohy, M., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Xu, C.N.; Zhang, H.J.; Zhou, Z.S.; Hu, T.; Wang, S.T.; Wang, Y.N.; Cao, K.Q. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Y.; He, W.; Zhang, Y. Stem blight of blueberry caused by Lasiodiplodia vaccinii sp. Nov. in China. Plant Dis. 2019, 103, 2041–2050. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Korf, I.; Robb, S.M.; Parra, G.; Ross, E.; Moore, B.; Holt, C.; Alvarado, A.S.; Yandell, M. MAKER: An easy-to-use annotation pipeline designed for emerging model organism genomes. Genome Res. 2008, 18, 188–196. [Google Scholar] [CrossRef]
- Todd, M.L.; Sean, R.E. tRNAscan-SE: A Program for Improved Detection of Transfer RNA Genes in Genomic Sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2013, 42, D699–D704. [Google Scholar] [CrossRef]
- Muller, J.; Szklarczyk, D.; Julien, P.; Letunic, I.; Roth, A.; Kuhn, M.; Powell, S.; von Mering, C.; Doerks, T.; Jensen, L.J. eggNOG v2. 0: Extending the evolutionary genealogy of genes with enhanced nonsupervised orthologous groups, species and functional annotations. Nucleic Acids Res. 2010, 38 (Suppl. 1), D190–D195. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39 (Suppl. 2), W29–W37. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef]
- Park, B.H.; Karpinets, T.V.; Syed, M.H.; Leuze, M.R.; Uberbacher, E.C. CAZymes Analysis Toolkit (CAT): Web service for searching and analyzing carbohydrate-active enzymes in a newly sequenced organism using CAZy database. Glycobiology 2010, 20, 1574–1584. [Google Scholar] [CrossRef]
- Urban, M.; Cuzick, A.; Seager, J.; Wood, V.; Rutherford, K.; Venkatesh, S.Y.; De Silva, N.; Martinez, M.C.; Pedro, H.; Yates, A.D.; et al. PHI-base: The pathogen-host interactions database. Nucleic Acids Res. 2019, 48, D613–D620. [Google Scholar] [CrossRef]
- Rafiei, V.; Velez, H.; Tzelepis, G. The Role of Glycoside Hydrolases in Phytopathogenic Fungi and Oomycetes Virulence. Int. J. Mol. Sci. 2021, 22, 9359. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant cell wall-degrading enzymes and their secretion in plant-pathogenic fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Thangavelu, B.; Chun, S.C.; Sathiyabama, M. Proteases from phytopathogenic fungi and their importance in phytopathogenicity. J. Gen. Plant Pathol. 2016, 82, 233–239. [Google Scholar] [CrossRef]
- Liu, J.; Wei, Y.; Yin, Y.; Zhu, K.; Liu, Y.; Ding, H.; Lei, J.; Zhu, W.; Zhou, Y. Effects of Mixed Decomposition of Pinus sylvestris var. mongolica and Morus alba Litter on Microbial Diversity. Microorganisms 2022, 10, 1117. [Google Scholar] [CrossRef]
- Liu, D.-M.; Huang, Y.-Y.; Liang, M.-H. Analysis of the probiotic characteristics and adaptability of Lactiplantibacillus plantarum DMDL 9010 to gastrointestinal environment by complete genome sequencing and corresponding phenotypes. LWT 2022, 158, 113129. [Google Scholar] [CrossRef]
- Bradley, E.L.; Ökmen, B.; Doehlemann, G.; Henrissat, B.; Bradshaw, R.E.; Mesarich, C.H. Secreted Glycoside Hydrolase Proteins as Effectors and Invasion Patterns of Plant-Associated Fungi and Oomycetes. Front. Plant Sci. 2022, 13, 853106. [Google Scholar] [CrossRef]
- Agrawal, Y.; Khatri, I.; Subramanian, S.; Shenoy, B.D. Genome sequence, comparative analysis, and evolutionary insights into chitinases of entomopathogenic fungus Hirsutella thompsonii. Genome Biol. Evol. 2015, 7, 916–930. [Google Scholar] [CrossRef]
- Crešnar, B.; Petrič, S. Cytochrome P450 enzymes in the fungal kingdom. Biochim. Biophys. Acta 2011, 1814, 29–35. [Google Scholar] [CrossRef]
- Mair, W.J.; Deng, W.; Mullins, J.G.; West, S.; Wang, P.; Besharat, N.; Ellwood, S.R.; Oliver, R.P.; Lopez-Ruiz, F.J. Demethylase inhibitor fungicide resistance in Pyrenophora teres f. sp. Teres associated with target site modification and inducible overexpression of CYP51. Front. Microbiol. 2016, 7, 1279. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Values | |
|---|---|---|
| Reads features (PacBio) | Total read number | 168,409 |
| Total bases (Mbp) | 3449.18 | |
| Average read length (Kbp) | 20.48 | |
| Largest read length (Kbp) | 55.65 | |
| Genome features | Predicted genome size (Mbp) | 48.36 |
| BUSCOs (%) | 98.6 | |
| GC content (%) | 51.73 | |
| Contig number | 8 | |
| Contig N50 (Mbp) | 6.62 | |
| Contig N90 (Mbp) | 5.65 | |
| Average contig length (Mbp) | 6.04 | |
| Largest contig length (Mbp) | 8.92 | |
| Repeat sequence (%) | 0.06 | |
| Protein-coding genes | 13,667 | |
| Gene Density (number of genes per Kbp) | 0.28 | |
| tRNA genes | 238 | |
| Candidate secreted proteins | 1580 | |
| Carbohydrate-active enzymes | 798 | |
| Pathogen host interactive genes | 2036 | |
| Fungal virulence factors | 1439 | |
| Gene ID | Location | PHI ID | Protein ID | Pathogen Species | Function | Identity (%) |
|---|---|---|---|---|---|---|
| gene00540 | Scaffold1 | PHI:4918 | B4EIW7 | Burkholderia cenocepacia | Global virulence regulator | 33.3 |
| gene00727 | Scaffold1 | PHI:7014 | Q63KU5 | Burkholderia pseudomallei | Putative peptide synthase/polyketide synthase; proteasome inhibitor | 23.9 |
| gene00747 | Scaffold1 | PHI:6957 | A0A0J9WX00 | Xanthomonas oryzae | Dehydrogenase | 25.6 |
| gene01771 | Scaffold1 | PHI:3607 | K7NCV2 | Epichloe festucae | non-ribosomal peptide synthetase Gene (sidN) encoding a siderophore synthetase | 35 |
| gene01838 | Scaffold1 | PHI:4613 | D0ZDL7 | Edwardsiella tarda | Type III Secretion System | 24.6 |
| gene02362 | Scaffold1 | PHI:3630 | L7N655 | Mycobacterium tuberculosis | Probable aminopeptidase | 34.1 |
| gene02365 | Scaffold1 | PHI:3793 | Q6FM27 | Candida glabrata | Encode alpha-(1-6)-mannosyl-transferases | 42.6 |
| gene02548 | Scaffold1 | PHI:2393 | I1R980 | Fusarium graminearum | - | 26.8 |
| gene02685 | Scaffold2 | PHI:2393 | I1R980 | Fusarium graminearum | - | 32.4 |
| gene02746 | Scaffold2 | PHI:7014 | Q63KU5 | Burkholderia pseudomallei | Putative peptide synthase/polyketide synthase; proteasome inhibitor | 33.6 |
| gene02944 | Scaffold2 | PHI:2393 | I1R980 | Fusarium graminearum | - | 28.7 |
| gene03052 | Scaffold2 | PHI:2313 | F8R4 × 8 | Metarhizium anisopliae | Dihydroxynaphthalene Melanin Biosynthesis | 67.1 |
| gene03080 | Scaffold2 | PHI:5268 | C8V0N8 | Aspergillus nidulans | Epimerase | 66.1 |
| gene03215 | Scaffold2 | PHI:2393 | I1R980 | Fusarium graminearum | - | 39.2 |
| gene03814 | Scaffold2 | PHI:7048 | Q5A663 | Candida albicans | Intramitochondrial quality control protease | 31.9 |
| gene03868 | Scaffold2 | PHI:2393 | I1R980 | Fusarium graminearum | - | 25.8 |
| gene03896 | Scaffold2 | PHI:3529 | J9VQL2 | Cryptococcus neoformans | Phosphate Acquisition and Storage | 46.9 |
| gene04733 | Scaffold3 | PHI:2393 | I1R980 | Fusarium graminearum | - | 26.8 |
| gene05524 | Scaffold3 | PHI:7121 | B0YCA2 | Aspergillus fumigatus | Trehalose–phosphate synthase subunit | 51.9 |
| gene05651 | Scaffold3 | PHI:2393 | I1R980 | Fusarium graminearum | - | 26 |
| gene05825 | Scaffold3 | PHI:9218 | B8NRT5 | Aspergillus flavus | Peptidyl–prolyl cis–trans isomerase | 40.4 |
| gene05939 | Scaffold3 | PHI:9236 | Q18CB9 | Clostridioides difficile | Cyclophilin-type peptidyl–prolyl-cis/trans–isomerase | 36 |
| gene06010 | Scaffold3 | PHI:9225 | A0A1D8PCN8 | Candida albicans | Phosphoprotein phosphatase PP4 regulatory subunit | 32.3 |
| gene06177 | Scaffold3 | PHI:9226 | A0A1D8PSJ8 | Candida albicans | Phosphoprotein phosphatase PP4 catalytic subunit | 68.2 |
| gene06187 | Scaffold3 | PHI:4468 | A0A068BFA5 | Epichloe festucae | Cell-Wall Integrity MAPK | 30.1 |
| gene06309 | Scaffold3 | PHI:7619 | Q8Y755 | Listeria monocytogenes | DExD-box RNA-helicase | 31.3 |
| gene06574 | Scaffold3 | PHI:3630 | L7N655 | Mycobacterium tuberculosis | Probable aminopeptidase | 30.2 |
| gene06896 | Scaffold4 | PHI:6751 | Q4WQ36 | Aspergillus fumigatus | Initiates asexual development | 38.1 |
| gene07319 | Scaffold4 | PHI:3085 | P42375 | Porphyromonas gingivalis | folding of newly synthesized proteins, preventing misfolding and aggregation | 53.6 |
| gene07390 | Scaffold4 | PHI:6548 | F9WZ47 | Zymoseptoria tritici | Involved in virulence and host-specific disease development | 32 |
| gene07781 | Scaffold4 | PHI:3630 | L7N655 | Mycobacterium tuberculosis | Probable aminopeptidase | 38.5 |
| gene08414 | Scaffold4 | PHI:494 | Q4WPX2 | Aspergillus fumigatus | Fatty acid oxygenase | 34.1 |
| gene08619 | Scaffold4 | PHI:5267 | E9R863 | Aspergillus nidulans | Epimerase | 52 |
| gene08738 | Scaffold5 | PHI:6956 | A0A0J9WWZ9 | Xanthomonas oryzae | 3-oxoacyl-ACP reductase | 28.7 |
| gene08982 | Scaffold5 | PHI:6956 | A0A0J9WWZ9 | Xanthomonas oryzae | 3-oxoacyl-ACP reductase | 28.2 |
| gene09589 | Scaffold5 | PHI:6751 | Q4WQ36 | Aspergillus fumigatus | Initiates asexual development | 29 |
| gene09837 | Scaffold5 | PHI:2393 | I1R980 | Fusarium graminearum | - | 28.9 |
| gene09987 | Scaffold5 | PHI:672 | P0CM56 | Cryptococcus neoformans | Capsule polysaccharide biosynthesis | 32.1 |
| gene10593 | Scaffold6 | PHI:2393 | I1R980 | Fusarium graminearum | - | 28.9 |
| gene10726 | Scaffold6 | PHI:6644 | J4KNV2 | Beauveria bassiana | Complex I intermediate-associated protein | 45.9 |
| gene10795 | Scaffold6 | PHI:7697 | A0A0F7A0T2 | Pseudomonas syringae | Red/far-red light-sensing bacteriophytochrome | 38 |
| gene10920 | Scaffold6 | PHI:494 | Q4WPX2 | Aspergillus fumigatus | Fatty acid oxygenase | 45.7 |
| gene11019 | Scaffold6 | PHI:5043 | E7Q5D7 | Saccharomyces cerevisiae | Hear shock protein | 49.5 |
| gene11058 | Scaffold6 | PHI:9526 | A0A384JQ57 | Botrytis cinerea | Pectin esterase | 48 |
| gene11108 | Scaffold6 | PHI:6751 | Q4WQ36 | Aspergillus fumigatus | Initiates asexual development | 30.7 |
| gene11239 | Scaffold6 | PHI:8509 | G9MQ09 | Trichoderma virens | Non-ribosomal peptide synthetase | 50.7 |
| gene11541 | Scaffold6 | PHI:2393 | I1R980 | Fusarium graminearum | - | 36.8 |
| gene11622 | Scaffold6 | PHI:7685 | A0A0J9UIM0 | Fusarium oxysporum | Nitrogen status-sensing regulatory protein | 53.1 |
| gene11701 | Scaffold6 | PHI:8896 | A0A1D8PKM5 | Candida albicans | Phosphatidyl-N-methylethanolamine N-methyltransferase | 59.5 |
| gene11796 | Scaffold6 | PHI:2383 | Q2VF46 | Monilinia fructicola | Cutinase | 35.6 |
| gene11972 | Scaffold6 | PHI:3417 | W5ZQ93 | Beauveria bassiana | Mitochondrial transmembrane protein | 36.3 |
| gene12319 | Scaffold7 | PHI:1134 | B0LLU1 | Leptosphaeria maculans | Unknown | 41.2 |
| gene12442 | Scaffold7 | PHI:9526 | A0A384JQ57 | Botrytis cinerea | Pectin esterase | 47.6 |
| gene12718 | Scaffold7 | PHI:8556 | O52658 | Pseudomonas aeruginosa | Acyl carrier protein | 37.8 |
| gene13268 | Scaffold7 | PHI:6090 | A0A0R1BRS8 | Acinetobacter nosocomialis | Hydroperoxidase | 45.8 |
| gene13405 | Scaffold7 | PHI:2393 | I1R980 | Fusarium graminearum | - | 34.5 |
| gene13409 | Scaffold7 | PHI:2393 | I1R980 | Fusarium graminearum | - | 24.2 |
| gene13412 | Scaffold7 | PHI:6956 | A0A0J9WWZ9 | Xanthomonas oryzae | 3-oxoacyl-ACP reductase | 25.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Wang, S.; Zhang, Y.; Chen, Y.; Zhou, H.; Zhang, G. Identification, Culture Characteristics and Whole-Genome Analysis of Pestalotiopsis neglecta Causing Black Spot Blight of Pinus sylvestris var. mongolica. J. Fungi 2023, 9, 564. https://doi.org/10.3390/jof9050564
Yang J, Wang S, Zhang Y, Chen Y, Zhou H, Zhang G. Identification, Culture Characteristics and Whole-Genome Analysis of Pestalotiopsis neglecta Causing Black Spot Blight of Pinus sylvestris var. mongolica. Journal of Fungi. 2023; 9(5):564. https://doi.org/10.3390/jof9050564
Chicago/Turabian StyleYang, Jing, Shuren Wang, Yundi Zhang, Yunze Chen, Heying Zhou, and Guocai Zhang. 2023. "Identification, Culture Characteristics and Whole-Genome Analysis of Pestalotiopsis neglecta Causing Black Spot Blight of Pinus sylvestris var. mongolica" Journal of Fungi 9, no. 5: 564. https://doi.org/10.3390/jof9050564