
Races CYR34 and Suwon11-1 of Puccinia striiformis f. sp. tritici Played an Important Role in Causing the Stripe Rust Epidemic in Winter Wheat in Yili, Xinjiang, China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wheat Stripe Rust Surveillance and Sample Collection
2.2. Spore Multiplication and Preservation
2.3. Race Identification
3. Results
3.1. Virulence Factors and Their Frequencies in Each Epidemic Region
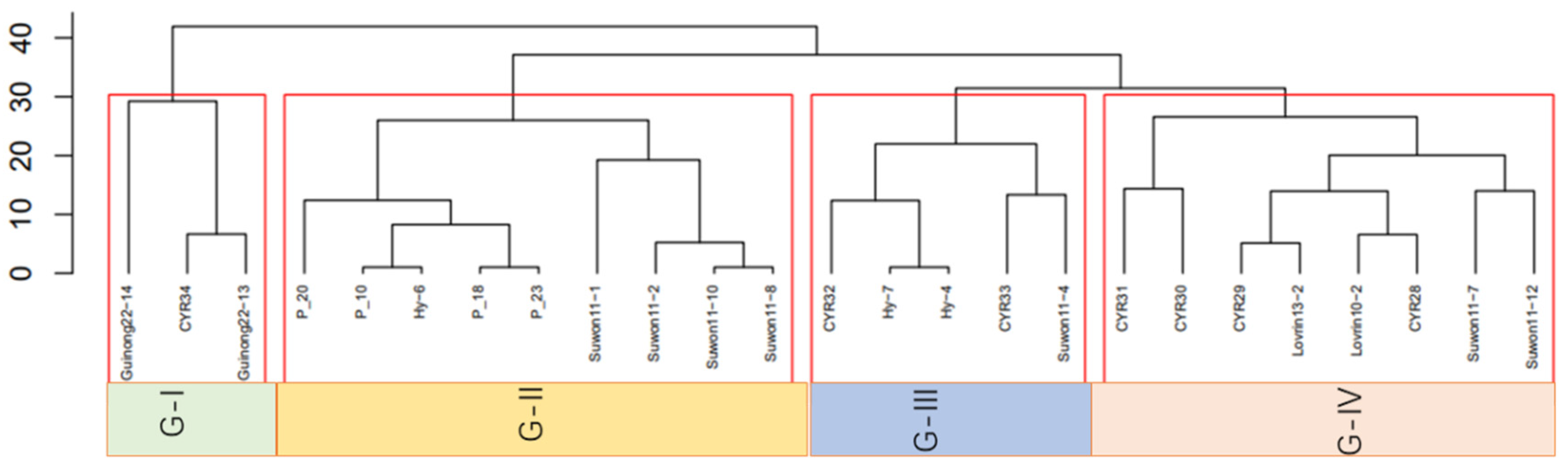
3.2. Race Groups through Cluster Analysis
3.3. Races in Different Epidemic Regions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biffen, R.H. Mendel’s laws of inheritance and wheat breeding. J. Agric. Sci. 1905, 1, 4–48. [Google Scholar] [CrossRef] [Green Version]
- Flor, H.H. Inheritance of pathogenicity in melampsora lini. Phytopathology 1942, 32, 653–669. [Google Scholar]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant cell wall-degrading enzymes and their secretion in plant-pathogenic fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.K.; Sanchez, A.; Angeles, E.; Singh, S.; Domingo, J.; Huang, N.; Khush, G.S. Are the dominant and recessive plant disease resistance genes similar? A case study of rice R genes and Xanthomonas oryzae pv. oryzae races. Genetics 2001, 159, 757–765. [Google Scholar] [CrossRef]
- Mundt, C.C. Pyramiding for resistance durability: Theory and practice. Phytopathology 2018, 108, 792–802. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Penman, L. Stripe rust epidemic and races of Puccinia striiformis in the United States in 2004. Phytopathology 2005, 95, S19. [Google Scholar]
- Lu, N.; Wang, J.; Chen, X.; Zhan, G.; Chen, C.; Huang, L.; Kang, Z. Spatial genetic diversity and interregional spread of Puccinia striiformis f. sp. tritici in Northwest China. Eur. J. Plant Pathol. 2011, 131, 685–693. [Google Scholar] [CrossRef]
- Zhan, G.; Ji, F.; Chen, X.; Wang, J.; Zhang, D.; Zhao, J.; Zeng, Q.; Yang, L.; Huang, L.; Kang, Z. Populations of Puccinia striiformis f. sp. tritici in Winter Spore Production Regions Spread from Southwestern Oversummering Areas in China. Plant Dis. 2022, 106, 2856–2865. [Google Scholar] [CrossRef]
- Fang, C.T. Physiologic specialization of Puccinia glumarum Erikss. and Henn. in China. Phytopathology 1944, 34, 2010–2014. [Google Scholar]
- Zhao, J.; Kang, Z. Fighting wheat rusts in China: A look back and into the future. Phytopathol. Res. 2023, 5, 6. [Google Scholar] [CrossRef]
- He, Z.H.; Rajaram, S.; Xin, Z.Y.; Huang, G.Z. A History of Wheat Breeding in China; International Maize and Wheat Improvement Center: DF (Mexico), Mexico, 2001. [Google Scholar]
- McIntosh, R.; Mu, J.; Han, D.; Kang, Z. Wheat stripe rust resistance gene Yr24/Yr26: A retrospective review. Crop J. 2018, 6, 321–329. [Google Scholar] [CrossRef]
- Wan, A.; Chen, X.; He, Z. Wheat stripe rust in China. Aust. J. Agric. Res. 2007, 58, 605–619. [Google Scholar] [CrossRef]
- Zhan, G.; Wang, F.; Wan, C.; Han, Q.; Huang, L.; Kang, Z.; Chen, X. Virulence and Molecular Diversity of the Puccinia striiformis f. sp. tritici Population in Xinjiang in Relation to Other Regions of Western China. Plant Dis. 2016, 100, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Liu, T.G.; Zhang, Z.Y.; Jia, Q.Z.; Wang, B.T.; Gao, L.; Peng, Y.L.; Jin, S.L.; Chen, W.Q. Discovery and pathogenicity of CYR34, a new race of Puccinia striiformis f. sp. tritici in China. Acta Phytopathol. Sin. 2017, 47, 681–687. [Google Scholar]
- Zhao, J.; Zhao, S.L.; Peng, Y.L.; Qin, J.F.; Huang, L.L.; Kang, Z.S. Investigation on geographic distribution and identification of six Berberis spp. serving as alternate host for Puccinia striiformis f. sp. tritici in Linzhi, Tibet. Acta Phytopathol. Sin. 2016, 46, 103–111. [Google Scholar]
- Garnica, D.P.; Nemri, A.; Upadhyaya, N.M.; Rathjen, J.P.; Dodds, P.N. The ins and outs of rust haustoria. PLoS Pathog. 2014, 10, e1004329. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Gladieux, P.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmoller, M.S.; Enjalbert, J.; De Vallavieille-Pope, C. Origin, migration routes and worldwide population genetic structure of the wheat yellow rust pathogen Puccinia striiformis f. sp. tritici. PLoS Pathog. 2014, 10, e1003903. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.M.; Yao, Q.; Huang, S.J.; Yan, J.H.; Hou, L.; Guo, Q.Y.; Zhao, J.; Kang, Z.S. Investigation and identification of barberry as alternate hosts for Puccinia striiformis f. sp. tritici in eastern Qinghai. Acta Phytopathol. Sin. 2019, 49, 370–378. [Google Scholar]
- Li, S.; Chen, W.; Ma, X.; Tian, X.; Liu, Y.; Huang, L.; Zhao, J. Identification of eight Berberis species from the Yunnan-Guizhou plateau as aecial hosts for Puccinia striiformis f. sp. Tritici, the wheat stripe rust pathogen. J. Integr. Agric. 2021, 20, 1563–1569. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, L.; Wang, Z.Y.; Chen, X.M.; Zhang, H.C.; Yao, J.N.; Zhan, G.M.; Chen, W.; Huang, L.L.; Kang, Z.S. Identification of eighteen Berberis species as alternate hosts of Puccinia striiformis f. sp. tritici and virulence variation in the pathogen isolates from natural infection of barberry plants in China. Phytopathology 2013, 103, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S.M.; Luo, Y. Long-distance spread and interregional epidemics of wheat stripe rust in China. Plant Dis. 2006, 90, 980–988. [Google Scholar] [CrossRef] [Green Version]
- Awais, M.; Ali, S.; Ju, M.; Liu, W.; Zhang, G.; Zhang, Z.; Li, Z.; Ma, X.; Wang, L.; Du, Z.; et al. Countrywide inter-epidemic region migration pattern suggests the role of southwestern population in wheat stripe rust epidemics in China. Environ. Microbiol. 2022, 24, 4684–4701. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Zeng, S.M. Wheat Rusts in China; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Wan, A.; Zhao, Z.; Chen, X.; He, Z.; Jin, S.; Jia, Q.; Yao, G.; Yang, J.; Wang, B.; Li, G.; et al. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [PubMed] [Green Version]
- Zhuang, H.; Zhao, J.; Huang, L.; Kang, Z.; Zhao, J. Identification of three Berberis species as potential alternate hosts for Puccinia striiformis f. sp. tritici in wheat-growing regions of Xinjiang, China. J. Integr. Agric. 2019, 18, 2786–2792. [Google Scholar] [CrossRef]
- Ali, S.; Gladieux, P.; Rahman, H.; Saqib, M.S.; Fiaz, M.; Ahmad, H.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmøller, M.S.; et al. Inferring the contribution of sexual reproduction, migration and off-season survival to the temporal maintenance of microbial populations: A case study on the wheat fungal pathogen Puccinia striiformis f. sp. tritici. Mol. Ecol. 2014, 23, 603–617. [Google Scholar] [CrossRef]
- Awais, M.; Zhao, J.; Cheng, X.; Khoso, A.G.; Ju, M.; Rehman, Z.U.; Iqbal, A.; Khan, M.R.; Chen, W.; Liu, M.; et al. Himalayan mountains imposing a barrier on gene flow of wheat yellow rust pathogen in the bordering regions of Pakistan and China. Fungal Genet. Biol. 2023, 164, 103753. [Google Scholar] [CrossRef]
- Ali, S.; Hodson, D. Wheat rust surveillance; field disease scoring and sample collection for phenotyping and molecular genotyping. In Methods in Molecular Biology; Periyannan, S., Ed.; Humana Press: New York, USA, 2017. [Google Scholar]
- Mehmood, S.; Sajid, M.; Husnain, S.K.; Zhao, J.; Huang, L.; Kang, Z. Study of Inheritance and Linkage of Virulence Genes in a Selfing Population of a Pakistani Dominant Race of Puccinia striiformis f. sp. tritici. Int. J. Mol. Sci. 2020, 21, 1685. [Google Scholar] [CrossRef] [Green Version]
- De Vallavieille-Pope, C.; Huber, L.; Leconte, M.; Bethenod, O. Preinoculation effects of light quantity on infection efficiency of Puccinia striiformis and P. triticina on wheat seedlings. Phytopathology 2002, 92, 1308–1314. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Chen, W.Q.; Wu, L.R.; Liu, T.G.; Xu, S.C.; Jin, S.L.; Peng, Y.L.; Wang, B.T. Race dynamics, diversity, and virulence evolution in Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust in China from 2003 to 2007. Plant Dis. 2009, 93, 1093–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Hao, Y.; Mergoum, M.; Bai, G.; Humphreys, G.; Cloutier, S.; Xia, X.; He, Z. Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- Wang, F.P.; Zhan, G.M.; Wei, G.R.; Huang, L.L.; Kang, Z.S.; Han, Q.M. Population virulence analysis of Puccinia striiformis f. sp. tritici on two wheat cultivars in differnet zones in longnan. J. Trit. Crops 2014, 34, 1146–1152. [Google Scholar]
- Sharma-Poudyal, D.; Chen, X.M.; Wan, A.M.; Zhan, G.M.; Kang, Z.S.; Cao, S.Q.; Jin, S.L.; Morgounov, A.; Akin, B.; Mert, Z.; et al. Virulence characterization of international collections of the wheat stripe rust pathogen, Puccinia striiformis f. sp. tritici. Plant Dis. 2013, 97, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lin, R.; Hu, J.; Shi, X.; Qiu, D.; Wu, P.; Goitom, G.H.; Wang, S.; Zhang, H.; Yang, L.; et al. Mapping of wheat stripe rust resistance gene Yr041133 by BSR-Seq analysis. Crop J. 2022, 10, 447–455. [Google Scholar] [CrossRef]
- Bai, B.; Du, J.Y.; Lu, Q.L.; He, C.Y.; Zhang, L.J.; Zhou, G.; Xia, X.C.; He, Z.H.; Wang, C.S. Effective resistance to wheat stripe rust in a region with high disease pressure. Plant Dis. 2014, 98, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Lan, C.X.; Singh, R.P.; Huerta-Espino, J.; Calvo-Salazar, V.; Herrera-Foessel, S.A. Genetic analysis of resistance to leaf rust and stripe rust in wheat cultivar Francolin#1. Plant Dis. 2014, 98, 1227–1234. [Google Scholar]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2017 Supplement; Yokohama, Japan Komugi Wheat Genetic Resources Database: Yokohama, Japan, 2017. [Google Scholar]
- Li, J.; Dundas, I.; Dong, C.; Li, G.; Trethowan, R.; Yang, Z.; Hoxha, S.; Zhang, P. Identification and characterization of a new stripe rust resistance gene Yr83 on rye chromosome 6R in wheat. Theor. Appl. Genet. 2020, 133, 1095–1107. [Google Scholar] [CrossRef]
- Wang, L.; Tang, X.; Wu, J.; Shen, C.; Dai, M.; Wang, Q.; Zeng, Q.; Kang, Z.; Wu, Y.; Han, D. Stripe rust resistance to a burgeoning Puccinia striiformis f. sp. tritici race CYR34 in current Chinese wheat cultivars for breeding and research. Euphytica 2019, 215, 68. [Google Scholar] [CrossRef]
- CNWRCG. Brief summary on identification of physiologic races of Puccinia striiformis f. sp. tritici in China in 1985 and 1986. Plant Prot. 1987, 13, 25–26. (In Chinese) [Google Scholar]
- Wang, F.L.; Wu, L.R.; Xu, S.C.; Jin, S.L.; Jia, Q.Z.; Yuan, W.H.; Yang, J.X. The discovery and studies on new races CYR30 and CYR31 of wheat stripe rust in China. Acta Phytopathol. Sin. 1996, 23, 39–44. (In Chinese) [Google Scholar]
- Wan, A.M.; Niu, Y.C.; Wu, L.R.; Yuan, W.H.; Li, G.B.; Jia, Q.Z.; Jin, S.L.; Yang, J.X.; Li, Y.F.; Bi, Y.Q. Physiologic specialization of stripe rust of wheat in China during 1991–1996. Acta Phytopathol. Sin. 1999, 29, 15–21. (In Chinese) [Google Scholar]
- Zhang, G.S.; Zhao, Y.Y.; Kang, Z.S.; Zhao, J. First report of a Puccinia striiformis f. sp. tritici race virulent to what stripe rust resistance gene Yr5 in China. Plant Dis. 2020, 104, 284. [Google Scholar] [CrossRef]
- Zhang, G.S.; Liu, W.; Wang, L.; Cheng, X.R.; Tian, X.X.; Du, Z.M.; Kang, Z.S.; Zhao, J. Evaluation on potential risk of the emerging Yr5-virulent races of Puccinia striiformis f. sp. tritici to 165 Chinese wheat cultivars. Plant Dis. 2022, 106, 1867–1874. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Gultyaeva, E.; Shaydayuk, E.; Kosman, E. Virulence Diversity of Puccinia striiformis f. sp. Tritici in Common Wheat in Russian Regions in 2019–2021. Agriculture 2022, 12, 1957. [Google Scholar] [CrossRef]
- Kang, Z.S.; Li, Z.Q. Discovery of a normal T. type new pathogenic strain to Lovrin 10. J. Northwest A. F. Univ. (Nat. Sci. Ed.) 1984, 4, 18–28. (In Chinese) [Google Scholar]
- Kang, Z.S.; Li, Z.Q.; Zhang, S.C. A preliminary studies of pathogenic strain to Lovrin 13 of yellow rust of wheat. J. Northwest A. F. Univ. (Nat. Sci. Ed.) 1987, 15, 105–106. (In Chinese) [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Differential Order | Virulence | Differential Lines | Yr Gene a |
|---|---|---|---|
| 1 | Vr1 | Trigo-Eureka | Yr6 |
| 2 | Vr2 | Fulhard | Unknown |
| 3 | Vr3 | Lutescens 128 | Unknown |
| 4 | Vr4 | Mentana | Unknown |
| 5 | Vr5 | Virgilio | YrVir1, YrVir2 |
| 6 | Vr6 | Abbondanza | Unknown |
| 7 | Vr7 | Early Premium | Unknown |
| 8 | Vr8 | Funo | YrA,+ |
| 9 | Vr9 | Danish 1 | Yr3 |
| 10 | Vr10 | JubilejinaII | YrJu1, YrJu2, YrJu3, YrJu4 |
| 11 | Vr11 | Fengchan 3 | Yr1 |
| 12 | Vr12 | Lovrin 13 | Yr9,+ |
| 13 | Vr13 | Kangyin 655 | Yr1, YrKy1, YrKy2 |
| 14 | Vr14 | Suwon 11 | YrSu |
| 15 | Vr15 | Zhong 4 | Unknown |
| 16 | Vr16 | Lovrin 10 | Yr9 |
| 17 | Vr17 | Hybrid 46 | Yr3b, Yr4b |
| 18 | Vr18 | Triticum spelta var. Album | Yr5 |
| 19 | Vr19 | Guinong22 | Yr10 , Yr26 |
| Virulence | Nileke | Gongliu | Huocheng | Qapqal | Xinyuan | Overall Population |
|---|---|---|---|---|---|---|
| Vr1 | 66.7 | 82.9 | 100.0 | 70.4 | 84.2 | 80.6 |
| Vr2 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Vr3 | 100.0 | 78.0 | 0.0 | 77.8 | 86.0 | 81.4 |
| Vr4 | 66.7 | 70.7 | 0.0 | 63.0 | 78.9 | 72.1 |
| Vr5 | 66.7 | 58.5 | 0.0 | 51.9 | 75.4 | 64.3 |
| Vr6 | 100.0 | 78.0 | 0.0 | 77.8 | 86.0 | 81.4 |
| Vr7 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Vr8 | 100.0 | 75.6 | 0.0 | 77.8 | 84.2 | 79.8 |
| Vr9 | 100.0 | 75.6 | 0.0 | 77.8 | 86.0 | 80.6 |
| Vr10 | 66.7 | 19.5 | 0.0 | 25.9 | 42.1 | 31.8 |
| Vr11 | 100.0 | 78.0 | 0.0 | 77.8 | 86.0 | 81.4 |
| Vr12 | 66.7 | 26.8 | 0.0 | 33.3 | 52.6 | 40.3 |
| Vr13 | 66.7 | 19.5 | 0.0 | 25.9 | 42.1 | 31.8 |
| Vr14 | 100.0 | 92.7 | 100.0 | 81.5 | 87.7 | 88.4 |
| Vr15 | 0.0 | 2.4 | 0.0 | 0.0 | 3.5 | 2.3 |
| Vr16 | 100.0 | 34.1 | 0.0 | 59.3 | 61.4 | 52.7 |
| Vr17 | 66.7 | 34.1 | 0.0 | 40.7 | 59.6 | 47.3 |
| Vr18 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Vr19 | 33.3 | 7.3 | 0.0 | 14.8 | 28.1 | 18.6 |
| Sample size | 3 | 41 | 1 | 27 | 57 | 129 |
| Virulence | Sample Size | Number of Races Detected | Race Diversity | Frequency (%) of the Most Frequent Race | Number of Virulences Detected | Frequency (%) of the Most Frequent Virulence |
|---|---|---|---|---|---|---|
| Nileke | 3 | 3 | 0.67 | 33.33 | 17 | 100 |
| Gongliu | 41 | 16 | 0.88 | 21.95 | 18 | 100 |
| Huocheng | 1 | 1 | 0.00 | 100.00 | 4 | 100 |
| Qapqal | 27 | 16 | 0.91 | 22.22 | 17 | 100 |
| Xinyuan | 57 | 17 | 0.89 | 22.81 | 18 | 100 |
| Overall population | 129 | 25 | 0.91 | 18.60 | 18 | 100 |
| Race | Virulence Profile a | Nileke | Gongliu | Huocheng | Qapqal | Xinyuan | Overall Population |
|---|---|---|---|---|---|---|---|
| CYR28 | 1,2,3,4,5,6,7,8,9,-,11,-,-,-,-,16,-,-,- | - | 2.4 | - | 7.4 | 3.5 | 3.9 |
| CYR29 | 1,2,3,4,5,6,7,8,9,-,11,12,-,-,-,16,-,-,- | - | - | - | 3.7 | - | 0.8 |
| CYR30 | 1,2,3,4,5,6,7,8,9,-,11,12,-,-,-,16,17,-,- | - | 2.4 | - | - | 7.0 | 3.9 |
| CYR31 | 1,2,3,4,5,6,7,8,9,-,11,12,-,14,-,16,17,-,- | - | 4.9 | - | 7.4 | 5.3 | 5.4 |
| CYR32 | 1,2,3,4,5,6,7,8,9,10,11,12,13,14,-,16,17,-,- | 33.3 | 12.2 | - | - | 8.8 | 8.5 |
| CYR33 | -,2,3,4,5,6,7,8,9,10,11,12,13,14,-,16,-,-,- | - | - | - | - | 1.8 | 0.8 |
| CYR34 | 1,2,3,4,5,6,7,8,9,10,11,12,13,14,-,16,17,-,19 | 33.3 | 4.9 | - | 7.4 | 22.8 | 14.0 |
| Guinong22-13 | -,2,3,-,-,6,7,8,9,10,11,12,13,14,-,16,17,-,19 | - | - | - | 7.4 | 1.8 | 2.3 |
| Guinong22-14 | 1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,-,-,-,19 | - | 2.4 | - | - | 3.5 | 2.3 |
| Hy-4 | 1,2,3,4,5,6,7,8,9,10,11,-,13,14,-,16,17,-,- | - | - | - | - | 3.5 | 1.6 |
| Hy-6 | 1,2,3,4,5,6,7,8,9,-,11,-,-,14,-,-,17,-,- | - | 2.4 | - | 7.4 | 5.3 | 4.7 |
| Hy-7 | -,2,3,4,5,6,7,8,9,10,11,-,13,14,-,16,17,-,- | - | - | - | 7.4 | - | 1.6 |
| Lovrin10-2 | 1,2,3,-,-,6,7,8,9,-,11,-,-,-,-,16,-,-,- | - | 2.4 | - | 3.7 | - | 1.6 |
| Lovrin13-2 | 1,2,3,4,-,6,7,8,9,-,11,12,-,-,-,16,-,-,- | - | - | - | 3.7 | 1.8 | 1.6 |
| P_10 | -,2,3,4,5,6,7,8,9,-,11,-,-,14,-,-,17,-,- | - | 2.4 | - | 3.7 | 3.5 | 3.1 |
| P_18 | 1,2,3,4,5,6,7,-,9,-,11,-,-,14,-,-,17,-,- | - | 2.4 | - | - | - | 0.8 |
| P_20 | -,2,3,4,5,6,7,8,-,-,11,-,-,14,-,-,17,-,- | - | 2.4 | - | - | - | 0.8 |
| P_23 | -,2,3,4,5,6,7,-,9,-,11,-,-,14,-,-,17,-,- | - | - | - | - | 1.8 | 0.8 |
| Suwon11-1 | 1,2,-,-,-,-,7,-,-,-,-,-,-,14,-,-,-,-,- | - | 22.0 | 100 | 22.2 | 14.0 | 18.6 |
| Suwon11-10 | -,2,3,4,-,6,7,8,9,-,11,-,-,14,-,-,-,-,- | - | 7.3 | - | 3.7 | 1.8 | 3.9 |
| Suwon11-12 | -,2,3,-,-,6,7,8,9,-,11,-,-,14,-,16,-,-,- | 33.3 | 4.9 | - | 3.7 | 5.3 | 5.4 |
| Suwon11-2 | 1,2,3,4,5,6,7,8,9,-,11,-,-,14,-,-,-,-,- | - | 19.5 | - | 3.7 | 8.8 | 10.9 |
| Suwon11-4 | -,2,3,4,-,6,7,8,9,10,11,-,13,14,-,16,-,-,- | - | - | - | 3.7 | - | 0.8 |
| Suwon11-7 | 1,2,3,4,5,6,7,8,9,-,11,12,-,14,-,16,-,-,- | - | - | - | 3.7 | - | 0.8 |
| Suwon11-8 | 1,2,3,4,-,6,7,8,9,-,11,-,-,14,-,-,-,-,- | - | 4.9 | - | - | - | 1.6 |
| Sample size | 3 | 41 | 1 | 27 | 57 | 129 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Awais, M.; Yang, H.; Shen, Y.; Li, G.; Gao, H.; Ma, J. Races CYR34 and Suwon11-1 of Puccinia striiformis f. sp. tritici Played an Important Role in Causing the Stripe Rust Epidemic in Winter Wheat in Yili, Xinjiang, China. J. Fungi 2023, 9, 436. https://doi.org/10.3390/jof9040436
Chen L, Awais M, Yang H, Shen Y, Li G, Gao H, Ma J. Races CYR34 and Suwon11-1 of Puccinia striiformis f. sp. tritici Played an Important Role in Causing the Stripe Rust Epidemic in Winter Wheat in Yili, Xinjiang, China. Journal of Fungi. 2023; 9(4):436. https://doi.org/10.3390/jof9040436
Chicago/Turabian StyleChen, Li, Muhammad Awais, Hong Yang, Yuyang Shen, Guangkuo Li, Haifeng Gao, and Jinbiao Ma. 2023. "Races CYR34 and Suwon11-1 of Puccinia striiformis f. sp. tritici Played an Important Role in Causing the Stripe Rust Epidemic in Winter Wheat in Yili, Xinjiang, China" Journal of Fungi 9, no. 4: 436. https://doi.org/10.3390/jof9040436