
Didymella pinodella: An Important Pea Root Rot Pathogen in France to Watch Out For?

1
Section of Ecological Plant Protection, University of Kassel, 37213 Witzenhausen, Germany
2
Section of Organic Plant Breeding and Agrobiodiversity, University of Kassel, 37213 Witzenhausen, Germany
3
Plant Protection Department, Faculty of Agronomy, University of the Oriental Republic of Uruguay (UDELAR), Research Station Dr. M.A. Cassinoni, Ruta 3 K 363, Paysandú 60000, Uruguay
*
Author to whom correspondence should be addressed.
J. Fungi 2024, 10(1), 44; https://doi.org/10.3390/jof10010044
Submission received: 27 November 2023
/
Revised: 24 December 2023
/
Accepted: 2 January 2024
/
Published: 5 January 2024
(This article belongs to the Section Fungal Pathogenesis and Disease Control)
Abstract
:Root rot pathogens restrict pea and wheat production globally. In the EU, pea and pea-based cereal mixtures are being promoted; however, root rot pathogen dynamics in such mixtures are poorly understood. Winter pea and wheat were grown either in pure stands or in mixtures in the field in western France, and the severity of root rot in pea, wheat, and their mixtures, as well as the key pathogens associated with these crops, were assessed. Disease severity was moderate in pea and low in wheat, with no effect of sowing pattern. Didymella pinodella, a previously unreported pathogen in the pea–root rot complex in France, emerged as the most dominant pathogen in pea. It also occurred in low frequencies in wheat. Subsequent greenhouse aggressiveness tests showed that ten of the commonly grown pea cultivars in France lack resistance to D. pinodella. Among the Fusarium spp. isolated, F. avenaceum was the most frequent, occurring at similar frequencies in pea and wheat. In conclusion, D. pinodella may be an important pea root rot pathogen in France and there is a lack of resistance in the tested pea cultivars. In addition, F. avenaceum is a shared pathogen of wheat and pea.
1. Introduction
Foot and root rots of pea and wheat are of great importance worldwide. They are caused by a multitude of fungal pathogens that usually co-occur as disease complexes [1,2,3,4]. Symptoms can occur throughout the season and mainly include damping off, foot and root necrosis, often accompanied with vascular tissue discolouration, and stunting, often causing significant yield losses, especially in wet years [2,4,5].
Several pathogens have been identified as the causal agents of foot and root rots in pea and wheat, some of which are shared between the two crops. Among the most common shared pathogens are Rhizoctonia, Pythium, and Fusarium species, including F. avenaceum, F. culmorum, F. tricinctum, F. oxysporum, and F. graminearum [2,3,6,7,8]. The populations of pathogens, i.e., the presence or absence and relative abundance of individual species, involved in the disease complex vary with geographical region and climatic conditions [6,7,9,10,11,12,13]. In addition to generalist pathogens, there are several specialist pathogens, such as Aphanomyces euteiches in pea and other legumes [7,14] and Gaeumannomyces graminis var. tritici in wheat and other small-grain cereals, that can cause severe yield losses [5].
In pea, Didymella pinodella (syn. Phoma pinodella, Phoma medicaginis var. pinodella) is an important pathogen causing foot and root rots across Canada [10,15] and Australia [16,17,18], as well as in several European countries, including Germany, Denmark, and Sweden, where it has been recognised as the major component of the pea–root rot complex [6,8,19,20]. Previous reports have highlighted the aggressive nature of D. pinodella in pea [17,21,22,23] and demonstrated its capacity to asymptomatically colonise wheat roots, resulting in biomass reduction [22]. This fungal pathogen causes diseases in various other legume crops, including lentils, chickpeas, and faba beans [11,24]; however, to date, D. pinodella has not been reported in France as a root rot pathogen, where it has mostly been implicated as a minor part of the pea–Ascochyta blight complex, which affects the leaves, stems, and pods [25].
This study was initiated to investigate the effects of pea and wheat mixtures compared to their pure stands on the severity of root rot and to characterise the populations of Fusarium and Didymella species associated with the roots of these crops. Given the emergence of D. pinodella as the predominant pathogen in the pea–root rot complex, the study objectives were extended to assess its aggressiveness and evaluate the susceptibility of important French pea cultivars to this pathogen.
2. Materials and Methods
2.1. Field Experiment
A full factorial experiment was conducted in a randomised complete block design with three replicates in the growing season of 2018/2019 at the experimental station of the France National Research Institute for Agriculture, Food and Environment (INRAE)—Institute for Genetics, Environment and Plant Protection (IGEPP), Rennes, western France. The experiment included the winter wheat cultivar ‘Cellule’ and the winter pea cultivar ‘Aviron’, grown either in pure stands, together as random mixtures, or in alternate rows. In mixed plantings, wheat was sown at 30% of the standard rate of 350–400 seeds m−2, whereas pea was sown at the recommended rate of 100% (90 seeds/m2). Seeds were planted at a depth of 3–4 cm in mid-October in 3 × 6 m plots, with a row spacing of 25 cm. The field experiment was initially planned for three consecutive growing seasons; however, sample collection was possible only in 2018/19. In the following season (2019/20), the experiment failed due to unfavourable weather conditions, and, in the 2020/21 sowing season, the experiment had to be cancelled due to the COVID-19 pandemic.
Foot and root rot disease severity was evaluated in 10 pea plants and 30 wheat tillers chosen randomly and dug up from each plot at the full flowering of pea. Pea root rot disease severity was visually assessed on a 0–8 scale (0 = healthy; 8 = dying plant) as described previously [12]. Disease severity classification for pea was as follows: no symptoms = plants with a score of 0, low = scores of 1–2, moderate = scores of 3–5, and high = scores of 6–8. The severity of foot rot symptoms in wheat was rated on a 0–3 scale according to Bockmann [26], with no symptoms = a score of 0; low = a score of 1, plants with lesions covering less than half of the stem circumference; moderate = a score of 2, plants with lesions spanning from 50% to 100% of the stem circumference; and high = a score of 3, plants with a rotten/broken stem [26]. There were no signs of any plants that were dying at the time of sampling.
2.2. Pathogen Isolations from Field Grown Plants and Morphlogical Identifications of Collected Isolates
A subset of six pea and six wheat plants from each plot were used for fungal isolation and morphological identification following the methods described in Šišić et al. [11]. Briefly, roots were surface sterilised for 10 s with 3% sodium hypochlorite, rinsed in distilled water and placed on filter paper under a laminar flow hood to dry. Three approximately 1 cm long pieces per plant, representing the root, crown, and transition zone, were placed on Coons’ agar [27]. Roots included lateral and tap roots up to the point of seed attachment. The crown was regarded as the point of seed attachment up to approximately 0.5 cm below the soil surface, and the transition zone included the next ca. 1.5 cm up towards the stem [11]. Plates were incubated for 2 weeks at 20 °C under 12 h cycles of blacklight blue fluorescent light and dark. Plates were examined for Didymella-like colonies, which were selected based on morphology and the production of pycnidia and/or the presence of chlamydospores [28]. Simultaneously, Fusarium-like colonies were selected based on colony morphology, pigmentation, and/or the presence of fusoid conidia [29]. Subsequently, both Didymella-like and Fusarium-like colonies were subcultured onto half-strength potato dextrose agar (19 g/L Difco PDA and 10 g/L agar Sigma Aldrich, Steinheim, Germany). Colonies were incubated for an additional 2–3 weeks and purified via the transfer of single pycnidia (Didymella morphology) or hyphal tipping (Fusarium morphology). Morphological identifications were then conducted according to the methods described by Boerema et al. [28] for Didymella isolates and Leslie and Summerell [29] for Fusarium isolates.
2.3. Molecular Validation of Fungal Species Identity and Phylogenetic Analyses
Three D. pinodella, four F. avenaceum, and nine Fusarium oxysporum species complex (FOSC) isolates were selected randomly and used to verify assigned morphological identity. Total genomic DNA was extracted from fresh mycelia collected from the fungal cultures actively growing on half-strength potato dextrose agar plates (19.5 g potato dextrose l-1 and 10 g agar l-1, Sigma Aldrich, Steinheim, Germany) following the protocol described by Sreelakshmi et al. [30]. The quantity and quality of DNAs were evaluated using a NanoDrop and stored in a TE buffer at −20 °C before PCR reactions.
The identity of D. pinodella isolates was confirmed by sequencing portions of the β tubulin (tub2) gene region with the primers Btub2Fd and Btub4Rd [31]. The identity of Fusarium spp. was verified through amplification and sequencing portions of the translation elongation factor 1α (EF-1α) gene region using the primer pairs EF1 and EF2 [32]. Amplicons for each locus were generated following the protocols described in Šišić et al. [33]. Amplicons were visualised via electrophoresis on a 1% agarose gel and purified using the DNA Clean & Concentrator kit (Zymo Research, Freiburg, Germany) according to the manufacturer’s instructions. Sanger sequencing was performed at Macrogen Europe Laboratories (Amsterdam, Netherlands) in both directions using the same primer pairs used for the PCR amplifications. Obtained row sequence data were assembled in SeqMan Lasergene software version 7.1.0 (DNAStar, Madison, WI, USA). The resulting consensus sequences were compared with the National Center for Biotechnology Information (NCBI) [34] and FUSARIUM-ID v.3.0 [35] databases. Further validation of the taxonomic assignments of the isolates was performed via phylogenetic analyses.
The reference sequences used for Didymella phylogeny were selected based on the Chen et al. 2017 [36] study. The reference sequences used for Fusarium phylogeny comprised one representative strain of each species complex in the F2 Fusarium clade (see Figure 1 in the Geiser et al., 2021 study [37]). Additional pea-root-associated FOSC reference sequences were included in the analysis based on recent pea root rot surveys conducted in France [38] and the UK [39]. Alignments were generated using a MAFFT sequence alignment server (https://mafft.cbrc.jp/alignment/server/, accessed on 20 December 2023) [40,41] and manually edited in MEGA v. 6.06. [42]. Phylogenetic inference was based on maximum likelihood (ML). The ML analyses were performed with the online version of the IQ-TREE software available at http://www.cibiv.at/software/iqtree, accessed on 20 December 2023 [43]. The ModelFinder option was used to identify the optimal partitioning scheme and substitution models. The branch support in IQ-TREE was completed by the Shimodaira–Hasegawa-like approximate likelihood ratio test (SH-aLRT) and the ultrafast bootstrap (UFBoot) with 1000 replicates. The sequences generated in this study, along with the reference sequences and their GenBank accession numbers, are listed in Tables S1 and S2.
2.4. Aggressiveness Tests in Pea
A greenhouse experiment was performed to test aggressiveness and to study the reaction of ten pea cultivars to D. pinodella isolates following methods previously described [22]. Of these, six cultivars were spring pea and four were winter pea, all provided by INRAE France and indicated as the most widely cultivated in France (Table 1).
Five D. pinodella isolates (FOEP 42.1500, FOEP 42.1501, FOEP 42.1503, FOEP 42.1027, and FOEP 42.1025) collected from symptomatic pea plants from the field experiment were grown on Coons’ agar for approximately 3 weeks under constant blacklight blue fluorescent light at 23 °C. Pycnidia and spores were collected from the agar surface using sterile microscope slides and approximately 15 mL of sterile distilled water. The suspension was passed through sterile cheesecloth to remove mycelial and pycnidial fragments, and spores were counted using a Fuchs Rosenthal hemocytometer (Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen, Germany).
Two surface-sterilised pea seeds (70% ethanol for 5 min) were planted per 300 mL pot that contained approximately 400 g of autoclaved sand. Following sowing, inoculations were carried out with either three single D. pinodella isolates (FOEP 42.1500, FOEP 42.1501, or FOEP 42.1503) or an equal mixture of the five isolates listed above. Inoculation was carried out by drenching the sand with spore suspensions to achieve a 2 × 104 spore g−1 substrate. Control pots were left non-inoculated and irrigated with distilled water. The experiment was arranged in a completely randomised design with four replicates for each pea cultivar. Pots were kept in the greenhouse at a 19 °C day and 16 °C night temperature, and a photoperiod of 16 h of light a day−1 (provided by 400 W high-pressure sodium lamps). Plants were watered daily with tap water. The number of surviving seedlings was recorded after 4 weeks. The plants were then removed from the pots, and the roots were separated from the above-ground parts, washed under running water, and evaluated for the severity of root rot symptoms and disease severity classes assigned as described above (Section 2.1). Above-ground fresh plant biomass was determined before drying at 105 °C until constant weight was achieved to determine dry plant biomass. In order to confirm that infection was a result of the inoculated pathogen, for each inoculated pea cultivar 4–6 roots were randomly selected, and D. pinodella isolates were re-isolated and identified morphologically using the protocol described above (Section 2.2.).
2.5. Data Analysis
All data were analysed using the statistical software R version 4.3.0 [45]. The analysis of ordinal (semi-quantitative) disease severity rating data from field and greenhouse experiments was performed using the Kruskal–Wallis test in the agricolae package [46]. Sowing pattern (field experiment), isolate, and genotype (greenhouse experiment) were considered fixed effects. If significant treatment effects were observed (p < 0.05), mean ranking values were separated with the Kruskal multiple comparison test [46,47]. The analysis of the sowing pattern effects on the isolation frequencies of individual fungal species recovered from pea roots from the field experiment was performed on proportional data using generalised linear models (bayesglm) with a binomial distribution and logit link function [48]. Contrasts (p < 0.05) were employed to separate the factor levels using the LSmeans package [49]. In the greenhouse inoculation experiment, numerous plants failed to emerge in the inoculated treatments. To account for this, prior to the data analysis, disease severity scores of 8 (corresponding to a dead plant) were manually added to the level of the cultivars’ corresponding non-inoculated control. This approach allowed for the calculation of the percent emergence for each cultivar tested, allowing for natural variation in seed germination among cultivars.
3. Results
3.1. Health Status of Field-Grown Plants
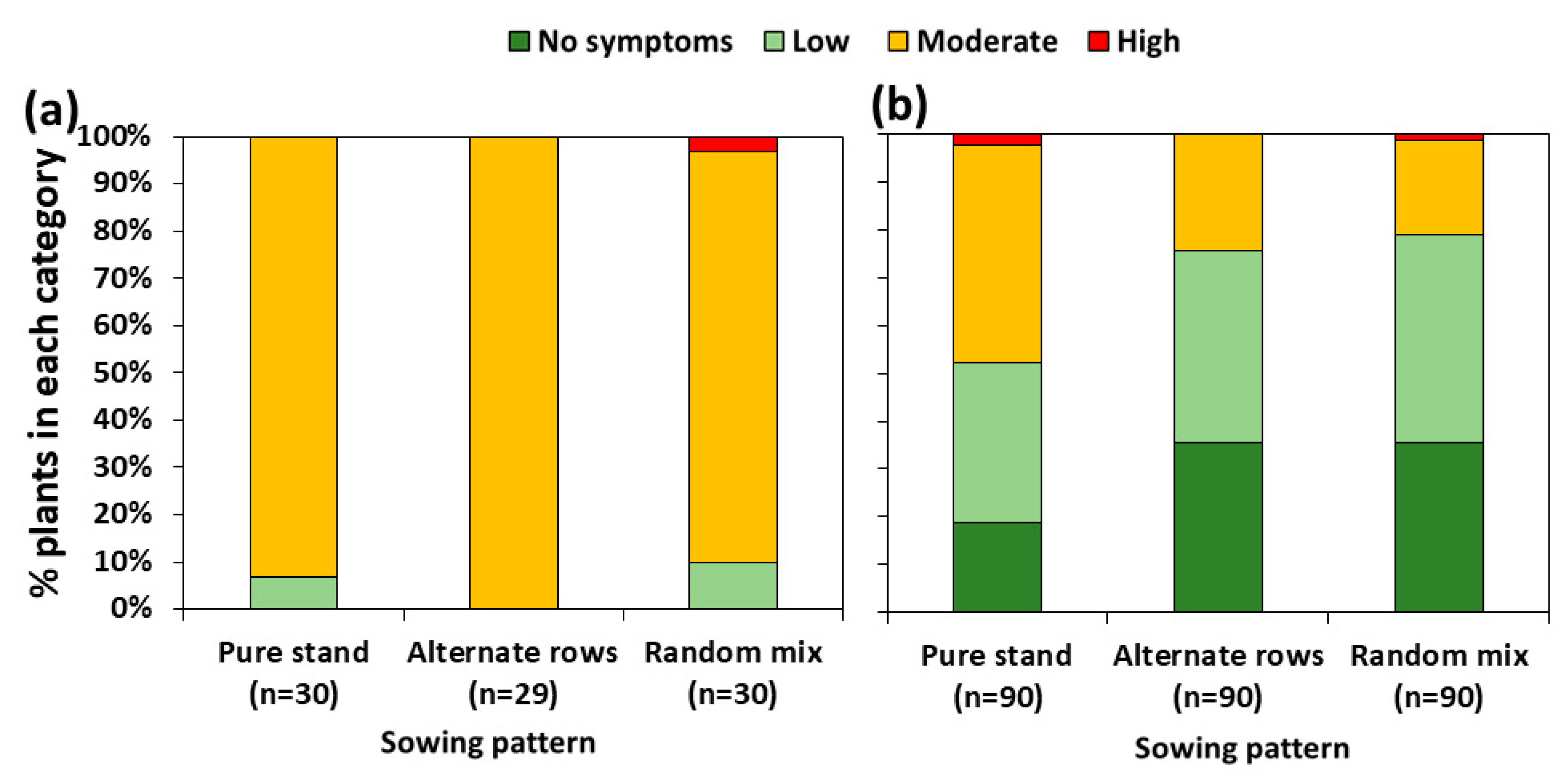
Almost all pea plants showed moderate levels of root rot symptom severity (Figure 1a), exhibiting brown to black necrotic lesions on stems and tap roots that totally encircled the tissue. About one-third of the wheat tillers assessed showed no symptoms of diseases (Figure 1b), whereas the remaining plants exhibited mostly streaks of reddish-brown discolourations on lower stems (low symptom severity), which sometimes expanded to form lesions encircling approximately 50% of the stem (moderate symptom severity). There were no significant differences in root rot symptom severity among sowing treatments (p = 0.7 for pea, and p = 0.2 for wheat) (Figure 1).
3.2. Composition of Fungal Pathogens Associated with the Roots of Field-Grown Plants
A total of 102 D. pinodella isolates were isolated from pea, where about 93% of the pea plants were infected by this pathogen, which represented about 58% of all of the isolates recovered from this host (Table 2). The pathogen was recovered at higher frequencies (p = 0.02) from pea stems (~85% infection rate) than roots (~41%) and at intermediate frequencies from crown tissue (~63%). Didymella pinodella also occurred in wheat, but with much lower isolation frequencies (approx. 15% of the plants were infected), accounting for about 13% of the isolates recovered from wheat where it was mainly recovered from crown tissue (Table 2). There was no effect of sowing pattern on D. pinodella isolation frequencies (Table 3).
Fusarium spp. were isolated at similar rates from both hosts, i.e., approx. 55% of the isolates originated from pea and approx. 45% isolates originated from wheat. In total, a total of 137 Fusarium isolates were recovered, representing six species, among which F. avenaceum was the most common, accounting for ~53% of all Fusarium isolates collected. This pathogen infected ~41% of the pea plants and ~39% of the wheat plants, and was predominantly isolated from the stem and crown tissue of both crops (20–35%) (Table 2). The members of the F. oxysporum (FOSC) and F. solani (FSSC) species complexes combined accounted for an additional ~36% of the Fusarium isolates. Both species complexes were common in pea (FOSC = ~35% and FSSC = ~22% of infected plants), but less frequent in wheat (FOSC = ~15% and FOSC = ~7% of infected plants). In pea, F. oxysporum was more frequently (p = 0.02) isolated from roots (approx. 22%) compared to stems (~4%). The type of plant tissue from which isolations were made had no significant effect on the isolation frequency of F. oxysporum from wheat (p = 0.8) nor F. solani from both crops (p > 0.3) (Table 2). Most remaining species found, including F. equiseti, F. crookwellense, F. dimerum, and D. pinodes, were represented by only a few isolates. As for D. pinodella, there was no effect of sowing pattern on the isolation frequencies of any of the Fusarium species (Table 3).
A phylogenetic analysis validated the morphologically assigned identifications of the isolates (Figure 2 and Figure 3). Didymella pinodella isolates matched D. pinodella CBS 531.66 and CBS 318.90 reference strains and were separated from the two sister species, D. pinodes and D. lethalis (Figure 2). Designated F. avenaceum isolates were placed in the Fusarium tricinctum species complex (FTSC), matching the F. avenaceum reference strain NRRL 54934. All FOSC isolates were accommodated in the F. oxysporum clade. A subgroup of eight isolates (three recovered from pea and five from wheat roots) closely matched the FOSC reference isolates MIAE08034, MIAE07954, PG57, and PG60, and two isolates (one recovered from pea and one from wheat roots) showed the closest genetic relationship with the F. oxysporum reference strains F233, PG108, and MIAI08036 (Figure 3). These reference isolates had previously been associated with diseased pea roots in France [26] and the UK [36].
3.3. Aggressiveness of Didymella pinodella to Pea in a Greenhouse
All three D. pinodella isolates, as well as the mixture of five isolates, were pathogenic to pea, causing different levels of pre-emergence damping off and root rot severity (Figure 4 and Figure 5a). In data analysed across pea cultivars (overall isolate effects), FOEP 42.1503 was classified as the most aggressive (p < 0.001), causing a 55% reduction in seedling emergence (n = 40/73, i.e., 33 inoculated plants emerged relative to a total of 73 plants which emerged in control treatments). Of the remaining FOEP 42.1503 inoculated pea plants (i.e., those that did emerge), approximately 88% (n = 29/33) developed severe root rot symptoms, displaying stunted growth, black lesions on the root system, and/or the collapse of the entire taproot. Only about 12% (n = 4/33) of the emerged plants that had been inoculated with FOEP 42.1503 developed moderate levels of root rot, displaying necrotic lesions on tap roots that completely encircled the root system mainly in the area around the seed attachment (Figure 4 and Figure 5a). The effects of D. pinodella isolates FOEP 42.1500 and FOEP 42.1501, including the mixture of five isolates, were similar in terms of aggressiveness. These treatments were somewhat lower in aggressiveness compared to the most aggressive isolate, FOEP 42.1503, but were also classified as highly aggressive, causing a 22% to 30% reduction in plant emergence, severe root rot on 18–39% of the emerged plants, and moderate levels of root rot symptom severity on 60–76% of the emerged plants (Figure 5a).
The pea cultivars (cvs.) used in this experiment showed some variability in their overall susceptibility, but these differences were not statistically significant (Figure 5b) (p = 0.4). Mean pre-emergence damping off ranged from 18% for cv. ‘Orchestra’ to 50% for cv. ‘Isard’ compared to the corresponding non-inoculated controls. Most of the remaining plants developed varying levels of root rot severity, ranging from moderate to high (Figure 5b). The reactions of individual cultivars indicated a wide range of susceptibility reactions and variations depending on the specific D. pinodella isolate used for inoculation (Figure 6). For example, FOEP 42.1503 caused an 83% reduction in plant emergence in the cv. ‘Poseidon’, and the effects of FOEP 42.1500 and the mixture of five isolates were intermediate (50% and 33% reduction in emergence, respectively), whereas the FOEP 42.1501 isolate was the least aggressive, causing no reduction in plant emergence and low to moderate root rot symptom severity in comparison to the corresponding non-inoculated control plants. By contrast, in the cv. ‘Safran’, isolates FOEP 42.1501 and FOEP 42.1503 caused severe root rot and pre-emergence damping off, the FOEP 42.1500 isolate effect was equally intermediate as for cv. ‘Poseidon’, and the mixture of five isolates was the least aggressive. Overall, none of the cultivars were resistant, with isolates causing varying levels of pre-emergence death and FOEP 42.1503 being consistently the most aggressive isolate in all pea cultivars (Figure 6).
4. Discussion
In the current study, the severity of foot and root rot symptoms in field-grown plants was moderate in pea and low to moderate in wheat. There was no effect of sowing pattern on the root rot symptom severity or the isolation frequencies of any of the fungal species. Didymella pinodella, a previously unreported pathogen in the pea–root rot complex in France, was the predominant pathogen in pea, recovered from 93% of symptomatic roots accounting for 58% of the isolates recovered from this host. On wheat, this pathogen occurred only sporadically. The greenhouse tests indicate the lack of resistance in ten widely grown French pea cultivars to D. pinodella and the potential of this pathogen to cause significant yield reductions. Fusarium spp. were isolated at moderate rates from both pea and wheat with F. avenaceum predominant. They occurred at similar frequencies on both hosts highlighting their significance as shared pathogens. Other, mostly specialised, pathogens, including F. oxysporum and F. solani, were mainly recovered from pea, whereas most of the remaining Fusarium species were represented by a few isolates only.
The emergence of D. pinodella at the research station in Rennes in western France as a predominant pathogen in the pea–root rot complex observed in our study on winter-grown peas contrasts the findings of a recent survey conducted in pea-growing areas across northern France on spring-grown green peas [38]. The authors reported that Fusarium spp. play a predominant role, whereas D. pinodella was not reported. One of the possible reasons for these contrasting results may be associated with the choice of agar media used to recover fungi from pea root pieces in the study of Gibert et al. [38]. As more than one single fungal species often colonise the roots simultaneously, the use of nutrient-rich malt extract agar for the culture-based fungal isolations, as employed by Gibert et al. [38], tends to favour fast-growing fungal species like Fusarium over slow-growing species such as D. pinodella [22,28]. We have found that using Coons’ agar [27] is particularly effective for isolating and identifying plant-associated D. pinodella. This medium promotes the growth of this pathogen and the formation of abundant pycnidia while also supporting the growth of Fusarium species. The agar choice as well as difficulties and often failure of cultural methods to recover other root pathogens, such as A. euteiches and F. sporotrichioides/F. culmorum in pea or Verticillium dahliae in chickpea, have been well documented [7,50,51,52]. It is also possible that the difference in the results between our study and Gibert et al. [38] could be attributed to site-specific agro-ecological effects, the type of pea, and/or the rotational history of the fields investigated. Gibert et al. [38] focused on spring green pea, which may have influenced the population of root-infecting fungi resulting in the reported prevalence of Fusarium species. Furthermore, in their study only 1 out of 22 sampled fields had a legume crop (i.e., common beans) in rotation 5 years prior to green pea sampling. Recent research has indicated, however, that an increase in D. pinodella abundance in the roots of pea and faba bean (Vicia faba) is linked to a greater frequency of these two [11,15,53] and likely other legume crops in rotation. The continued monitoring of this potentially important pea pathogen under field conditions is recommended.
The greenhouse data evaluating the resistance of widely grown pea cultivars in France, including both spring and winter varieties, to three D. pinodella isolates and the five-isolate mixture, indicated that all ten tested cultivars exhibited high susceptibility. Some variability in the reaction of individual pea cultivars to specific isolates was observed, with none of the cultivars showing high resistance to any of the isolates tested. All cultivars reacted with reduced emergence upon inoculation with at least one of the D. pinodella isolates. The emerged plants displayed moderate to severe root rot symptoms. Among individual isolates, the FOEP 42.1503 isolate was consistently the most aggressive. The symptoms caused by the remaining D. pinodella isolates, including the mixture of five isolates, were somewhat lower compared to the highly aggressive FOEP 42.1503 isolate and better corresponded to the symptoms observed in the field-grown plants. The ability of D. pinodella to cause severe root rot agrees with previous studies that also demonstrated the high pathogenicity of this pathogen to pea [22,54].
Taken together, the apparent dominance of D. pinodella in the pea–root rot complex observed in pea originating from northwestern France and the lack of resistance in tested pea cultivars warrants the need for additional research to monitor this pathogen in the major pea-growing areas of France. In order to overcome the limitations associated with culture-based fungal isolation, DNA-based detection techniques, such as quantitative real-time PCR assays, should be utilised [22,52,55].
The role of F. avenaceum in pea and wheat health is well documented. It is a serious and often highly aggressive pathogen in legumes and cereals [12,13,56,57], as well as a predominant species in the pea root rot complex in many regions of the world [6,7,58]. Our results suggest that F. avenaceum can become problematic in areas where legume and cereal crops are grown together in mixtures or in rotation as it can easily spread from one crop to the other. This pathogen may also play a role in the future expansion and development of important cereal disease complexes such as Fusarium head blight [59]. The members of the F. oxysporum and F. solani species complexes often show a high degree of host specificity, and both are well recognised as important pathogens of legumes but not cereals [11,29,57]. Both species complexes were reported as the most frequently detected in symptomatic spring pea root rots in 2017 across northern France [38]. The species F. equiseti has been previously shown to contribute to the reduction in diseases in various crops [60,61,62], including pea root rot caused by D. pinodella and F. avenaceum [21], the two most commonly isolated pathogens in this study.
5. Conclusions
This study brings new information about the root rot complex of pea and wheat in France. It highlights, on the one hand, that winter- and spring-grown peas may harbour quite different pathogen communities and substantial differences with respect to the importance of D. pinodella. On the other hand, it underscores the importance of F. avenaceum as a shared pathogen in both crops. This may be of concern for French pea production, given the highly aggressive nature of D. pinodella coupled with the lack of resistance in the 10 pea cultivars tested in the greenhouse. Further research is thus recommended to monitor this pathogen in major pea-growing regions of France. Utilising DNA-based detection techniques, particularly quantitative real-time PCR assays [22], would enhance the precision of such monitoring efforts. Furthermore, the assessment of D. pinodella resistance in a larger set of pea cultivars representing the wider pea gene pool is recommended.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof10010044/s1, Table S1: Genbank accession numbers for the β tubulin (tub2) gene region of the Didymella pinodella sequences generated in this study, along with the Didymella spp. reference strains and their GenBank accession numbers used to examine phylogenetic relationships among collected Didymella isolates. Table S2: Genbank accession numbers for the translation elongation factor 1α (EF-1α) gene region of the Fusarium sequences generated in this study, along with the Fusarium reference strains and their GenBank accession numbers used to examine phylogenetic relationships among collected Fusarium isolates.
Author Contributions
Conceptualization, A.Š.; methodology, A.Š. and J.B.-Š.; validation, all; formal analysis, A.Š.; investigation, A.Š., J.B.-Š. and F.M.G.; resources, A.Š. and M.R.F.; writing—original draft preparation, A.Š.; visualization, A.Š.; funding acquisition, A.Š. and M.R.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the EU project ReMIX, “Redesigning European cropping systems based on species MIXtures”, funded by the EU’s Horizon 2020 Research and Innovation Programme under grant 727217.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are contained within the article and Supplementary Materials.
Acknowledgments
The authors would like to express their gratitude to the members of the French National Institute for Agriculture, Food, and Environment (INRAE), specifically Alain Baranger, for coordinating the field experiment and sample collection, and to Angelique Lesne and Marie-Laure Pilet-Nayel for generously providing pea cultivars used in greenhouse aggressiveness tests.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dawkins, T.C.; Smiley, R.W.; Cook, R.J. Insights into the Prevalence and Management of Soilborne Cereal Pathogens under Direct Seeding in the Pacific Northwest, U.S.A. Can. J. Plant Pathol. 2002, 24, 416–428. [Google Scholar] [CrossRef]
- Parry, D.W. The Cereal Fusarium Complex. In Ecology of Plant Pathogens; Blakeman, J.P., Williamson, B., Eds.; CAB International: Wallingford, UK, 1994; Volume 4, pp. 301–320. [Google Scholar]
- Tillmann, M.; von Tiedemann, A.; Winter, M. Crop Rotation Effects on Incidence and Diversity of Fusarium Species Colonizing Stem Bases and Grains of Winter Wheat. J. Plant Dis. Prot. 2017, 124, 121–130. [Google Scholar] [CrossRef]
- Wille, L.; Messmer, M.M.; Studer, B.; Hohmann, P. Insights to Plant–Microbe Interactions Provide Opportunities to Improve Resistance Breeding against Root Diseases in Grain Legumes. Plant Cell Environ. 2019, 42, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Schoeny, A.; Jeuffroy, M.H.; Lucas, P. Influence of Take-All Epidemics on Winter Wheat Yield Formation and Yield Loss. Phytopathology 2001, 91, 694–701. [Google Scholar] [CrossRef]
- Baćanović-Šišić, J.; Šišić, A.; Schmidt, J.H.; Finckh, M.R. Identification and Characterization of Pathogens Associated with Root Rot of Winter Peas Grown under Organic Management in Germany. Eur. J. Plant Pathol. 2018, 151, 745–755. [Google Scholar] [CrossRef]
- Chatterton, S.; Harding, M.W.; Bowness, R.; Mclaren, D.L.; Banniza, S.; Gossen, B.D. Importance and Causal Agents of Root Rot on Field Pea and Lentil on the Canadian Prairies, 2014–2017. Can. J. Plant Pathol. 2019, 41, 98–114. [Google Scholar] [CrossRef]
- Šišić, A.; Baćanović-Šišić, J.; Schmidt, H.; Finckh, M.R. Root Pathogens Occurring on Pea (Pisum sativum) and Faba Bean (Vicia faba) in Germany. In 30th Scientific-Experts Conference of Agriculture and Food Industry: Answers for Forthcoming Challenges in Modern Agriculture; Brka, M., Omanović-Mikličanin, E., Karić, L., Falan, V., Toroman, A., Eds.; IFMBE Proceedings; Springer International Publishing: Cham, Switzerland, 2020; Volume 78, pp. 69–75. ISBN 978-3-030-40048-4. [Google Scholar]
- Bateman, G.L.; Murray, G. Seasonal Variations in Populations of Fusarium Species in Wheat-Field Soil. Appl. Soil Ecol. 2001, 18, 117–128. [Google Scholar] [CrossRef]
- Esmaeili Taheri, A.; Chatterton, S.; Foroud, N.A.; Gossen, B.D.; McLaren, D.L. Identification and Community Dynamics of Fungi Associated with Root, Crown, and Foot Rot of Field Pea in Western Canada. Eur. J. Plant Pathol. 2016, 147, 489–500. [Google Scholar] [CrossRef]
- Šišić, A.; Baćanović-Šišić, J.; Schmidt, H.; Finckh, M.R. Farming System Effects on Root Rot Pathogen Complex and Yield of Faba Bean (Vicia faba) in Germany. Front. Plant Sci. 2022, 13, 1009906. [Google Scholar] [CrossRef]
- Šišić, A.; Baćanović-Šišić, J.; Karlovsky, P.; Wittwer, R.; Walder, F.; Campiglia, E.; Radicetti, E.; Friberg, H.; Baresel, J.P.; Finckh, M.R. Roots of Symptom-Free Leguminous Cover Crop and Living Mulch Species Harbor Diverse Fusarium Communities That Show Highly Variable Aggressiveness on Pea (Pisum sativum). PLoS ONE 2018, 13, e0191969. [Google Scholar] [CrossRef]
- Uhlig, S.; Jestoi, M.; Parikka, P. Fusarium Avenaceum—The North European Situation. Int. J. Food Microbiol. 2007, 119, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Finckh, M.R.; Yli-Mattila, T.; Nykänen, A.; Kurki, P.; Hannukkala, A. Organic Temperate Legume Disease Management. In Plant Diseases and Their Management in Organic Agriculture; Finckh, M.R., van Bruggen, A.H.C., Tamm, L., Eds.; APS Press: Saint Paul, MN, USA, 2015; pp. 293–311. ISBN 978-0-89054-476-1. [Google Scholar]
- Bainard, L.D.; Navarro-Borrell, A.; Hamel, C.; Braun, K.; Hanson, K.; Gan, Y. Increasing the Frequency of Pulses in Crop Rotations Reduces Soil Fungal Diversity and Increases the Proportion of Fungal Pathotrophs in a Semiarid Agroecosystem. Agric. Ecosyst. Environ. 2017, 240, 206–214. [Google Scholar] [CrossRef]
- Bretag, T.W.; Keane, P.J.; Price, T.V. The Epidemiology and Control of Ascochyta Blight in Field Peas: A Review. Aust. J. Agric. Res. 2006, 57, 883. [Google Scholar] [CrossRef]
- Tran, H.S.; You, M.P.; Khan, T.N.; Barbetti, M.J. Pea Black Spot Disease Complex on Field Pea: Dissecting the Roles of the Different Pathogens in Causing Epicotyl and Root Disease. Eur. J. Plant Pathol. 2016, 144, 595–605. [Google Scholar] [CrossRef]
- Tran, H.S.; Li, Y.P.; You, M.P.; Khan, T.N.; Pritchard, I.; Barbetti, M.J. Temporal and Spatial Changes in the Pea Black Spot Disease Complex in Western Australia. Plant Dis. 2014, 98, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Persson, L.; Bødker, L.; Larsson-Wikström, M. Prevalence and Pathogenicity of Foot and Root Rot Pathogens of Pea in Southern Scandinavia. Plant Dis. 1997, 81, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Pflughöft, O.; Merker, C.; von Tiedemann, A.; Schäfer, B.C. Zur Verbreitung und Bedeutung von Pilzkrankheiten in Körnerfuttererbsen (Pisum sativum L.) in Deutschland. Gesunde Pflanz. 2012, 64, 39–48. [Google Scholar] [CrossRef]
- Šišić, A.; Baćanović, J.; Finckh, M.R. Endophytic Fusarium equiseti Stimulates Plant Growth and Reduces Root Rot Disease of Pea (Pisum sativum L.) Caused by Fusarium avenaceum and Peyronellaea pinodella. Eur. J. Plant Pathol. 2017, 148, 271–282. [Google Scholar] [CrossRef]
- Šišić, A.; Oberhänsli, T.; Baćanović-Šišić, J.; Hohmann, P.; Finckh, M.R. A Novel Real Time PCR Method for the Detection and Quantification of Didymella pinodella in Symptomatic and Asymptomatic Plant Hosts. J. Fungi 2022, 8, 41. [Google Scholar] [CrossRef]
- Schmidt, J.H.; Theisgen, L.V.; Finckh, M.R.; Šišić, A. Increased Resilience of Peas Toward Root Rot Pathogens Can Be Predicted by the Nematode Metabolic Footprint. Front. Sustain. Food Syst. 2022, 6, 881520. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. Available online: http://nt.ars-grin.gov/fungaldatabases/ (accessed on 3 July 2021).
- Onfroy, C.; Tivoli, B.; Corbière, R.; Bouznad, Z. Cultural, Molecular and Pathogenic Variability of Mycosphaerella pinodes and Phoma medicaginis var. pinodella Isolates from Dried Pea (Pisum sativum) in France. Plant Pathol. 1999, 48, 218–229. [Google Scholar] [CrossRef]
- Bockmann, H. Künstliche Freilandinfektionen Mit Den Erregern Der Fuß-Und Ährenkrankheiten Des Weizens. I. Vorbereitung Und Durchführung Der Feldinfektionen Sowie Deren Nebenwirkungen. Nachrichtenblatt Dtsch. Pflanzenschutz 1962, 10, 153–156. [Google Scholar]
- Coons, G.H. Factors Involved in the Growth and the Pycnidium Formation of Plenodomus fuscomaculans. J. Agric. Res. 1916, 5, 713–769. [Google Scholar]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004; ISBN 978-0-85199-743-8. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing: Ames, IA, USA, 2006; ISBN 978-0-8138-1919-8. [Google Scholar]
- Sreelakshmi, Y.; Gupta, S.; Bodanapu, R.; Chauhan, V.S.; Hanjabam, M.; Thomas, S.; Mohan, V.; Sharma, S.; Srinivasan, R.; Sharma, R. NEATTILL: A Simplified Procedure for Nucleic Acid Extraction from Arrayed Tissue for Tilling and Other High-Throughput Reverse Genetic Applications. Plant Methods 2010, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Aveskamp, M.M.; Woudenberg, J.H.C.; De Gruyter, J.; Turco, E.; Groenewald, J.Z.; Crous, P.W. Development of Taxon-Specific Sequence Characterized Amplified Region (SCAR) Markers Based on Actin Sequences and DNA Amplification Fingerprinting (DAF): A Case Study in the Phoma Exigua Species Complex. Mol. Plant Pathol. 2009, 10, 403–414. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple Evolutionary Origins of the Fungus Causing Panama Disease of Banana: Concordant Evidence from Nuclear and Mitochondrial Gene Genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Šišić, A.; Al-Hatmi, A.M.S.; Baćanović-Šišić, J.; Ahmed, S.A.; Dennenmoser, D.; de Hoog, G.S.; Finckh, M.R. Two New Species of the Fusarium solani Species Complex Isolated from Compost and Hibiscus (Hibiscus sp.). Antonie Van Leeuwenhoek 2018, 111, 1785–1805. [Google Scholar] [CrossRef]
- Torres-Cruz, T.J.; Whitaker, B.K.; Proctor, R.H.; Broders, K.; Laraba, I.; Kim, H.S.; Brown, D.W.; O’Donnell, K.; Estrada-Rodríguez, T.; Lee, Y.H.; et al. FUSARIUM-ID v.3.0: An Updated, Downloadable Resource for Fusarium Species Identification. Plant Dis. 2022, 106, 1610–1616. [Google Scholar] [CrossRef]
- Madden, T. The BLAST Sequence Analysis Tool. In The NCBI Handbook; McEntyre, J., Ostell, J., Eds.; National Center for Biotechnology Information: Bethesda, MD, USA, 2002; Chapter 16. [Google Scholar]
- Chen, Q.; Hou, L.W.; Duan, W.J.; Crous, P.W.; Cai, L. Didymellaceae Revisited. Stud. Mycol. 2017, 87, 105–159. [Google Scholar] [CrossRef]
- Geiser, D.M.; Al-Hatmi, A.M.S.; Aoki, T.; Arie, T.; Balmas, V.; Barnes, I.; Bergstrom, G.C.; Bhattacharyya, M.K.; Blomquist, C.L.; Bowden, R.L.; et al. Phylogenomic Analysis of a 55.1-Kb 19-Gene Dataset Resolves a Monophyletic Fusarium That Includes the Fusarium solani Species Complex. Phytopathology 2021, 111, 1064–1079. [Google Scholar] [CrossRef]
- Gibert, S.; Edel-Hermann, V.; Gautheron, E.; Gautheron, N.; Bernaud, E.; Sol, J.M.; Capelle, G.; Galland, R.; Bardon-Debats, A.; Lambert, C.; et al. Identification, Pathogenicity and Community Dynamics of Fungi and Oomycetes Associated with Pea Root Rot in Northern France. Plant Pathol. 2022, 71, 1550–1569. [Google Scholar] [CrossRef]
- Jenkins, S. Examining Identity, Phylogeny and Pathogenicity Factors in Fusarium Species Affecting Pea. Ph.D. Thesis, University of Warwick, Coventry, UK, 2018. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kuraku, S.; Zmasek, C.M.; Nishimura, O.; Katoh, K. aLeaves Facilitates On-Demand Exploration of Metazoan Gene Family Trees on MAFFT Sequence Alignment Server with Enhanced Interactivity. Nucleic Acids Res. 2013, 41, W22–W28. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Boutet, G.; Alves Carvalho, S.; Falque, M.; Peterlongo, P.; Lhuillier, E.; Bouchez, O.; Lavaud, C.; Pilet-Nayel, M.L.; Rivière, N.; Baranger, A. SNP Discovery and Genetic Mapping Using Genotyping by Sequencing of Whole Genome Genomic DNA from a Pea RIL Population. BMC Genom. 2016, 17, 121. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013.
- Mendiburu, D.F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-7. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 20 December 2023).
- Conover, W.J. Practical Nonparametric Statistics, 3rd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1999; ISBN 0-471-16068-7. [Google Scholar]
- Yu-Sung, S. Package ‘Arm’: Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Armstrong-Cho, C.; Sivachandra Kumar, N.T.; Kaur, R.; Banniza, S. The Chickpea Root Rot Complex in Saskatchewan, Canada-Detection of Emerging Pathogens and Their Relative Pathogenicity. Front. Plant Sci. 2023, 14, 1117788. [Google Scholar] [CrossRef]
- Armstrong-Cho, C.; Tetreault, M.; Banniza, S.; Bhadauria, V.; Morrall, R.A.A. First report of Aphanomyces euteiches in Saskatchewan. Can. Plant Dis. Surv. 2014, 94, 193–194. [Google Scholar]
- Zitnick-Anderson, K.; Simons, K.; Pasche, J.S. Detection and qPCR Quantification of Seven Fusarium Species Associated with the Root Rot Complex in Field Pea. Can. J. Plant Pathol. 2018, 40, 261–271. [Google Scholar] [CrossRef]
- Finckh, M.R.; Junge, S.M.; Schmidt, J.H.; Sisic, A.; Weedon, O.D. Intra- and Interspecific Diversity: The Cornerstones of Agroecological Crop Health Management. Asp. Appl. Biol. 2021, 146, 14. [Google Scholar]
- Šišić, A.; Baćanović-Šišić, J.; Finckh, M.R. Molecular Characterization and Aggressiveness of Didymella pinodella Isolates Associated with Root Rot of Field Pea (Pisum sativum). In Proceedings of the 61 Deutsche Pflanzenschutztagung “Herausforderung Pflanzenschutz—Wege in die Zukunft”, Universität Hohenheim, Stuttgart, Germany, 11–14 September 2018; p. 326. [Google Scholar]
- Elbelt, E.; Siou, D.; Gelisse, S.; Cruaud, C.; Lannou, C.; Lebrun Marc, H.; Laval, V. Optimized Real Time QPCR Assays for Detection and Quantification of Fusarium and Microdochium Species Involved in Wheat Head Blight as Defined by MIQE Guidelines. bioRxiv 2018, 272534. [Google Scholar] [CrossRef]
- Pettitt, T.; Xu, X.; Parry, D. Association of Fusarium Species in the Wheat Stem Rot Complex. Eur. J. Plant Pathol. 2003, 109, 769–774. [Google Scholar] [CrossRef]
- Safarieskandari, S.; Chatterton, S.; Hall, L.M. Pathogenicity and Host Range of Fusarium Species Associated with Pea Root Rot in Alberta, Canada. Can. J. Plant Pathol. 2020, 43, 162–171. [Google Scholar] [CrossRef]
- Chittem, K.; Mathew, F.M.; Gregoire, M.; Lamppa, R.S.; Chang, Y.W.; Markell, S.G.; Bradley, C.A.; Barasubiye, T.; Goswami, R.S. Identification and Characterization of Fusarium spp. Associated with Root Rots of Field Pea in North Dakota. Eur. J. Plant Pathol. 2015, 143, 641–649. [Google Scholar] [CrossRef]
- Osborne, L.E.; Stein, J.M. Epidemiology of Fusarium Head Blight on Small-Grain Cereals. Int. J. Food Microbiol. 2007, 119, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, H.; Muslim, A.; Suzuki, T.; Hyakumachi, M. Fusarium equiseti GF191 as an Effective Biocontrol Agent against Fusarium Crown and Root Rot of Tomato in Rock Wool Systems. Crop Prot. 2007, 26, 1514–1523. [Google Scholar] [CrossRef]
- Macia-Vicente, J.G.; Rosso, L.C.; Ciancio, A.; Jansson, H.B.; Lopez-Llorca, L.V. Colonisation of Barley Roots by Endophytic Fusarium equiseti and Pochonia chlamydosporia: Effects on Plant Growth and Disease. Ann. Appl. Biol. 2009, 155, 391–401. [Google Scholar] [CrossRef]
- Saldajeno, M.G.B.; Hyakumachi, M. The Plant Growth-Promoting Fungus Fusarium equiseti and the Arbuscular Mycorrhizal Fungus Glomus mosseae Stimulate Plant Growth and Reduce Severity of Anthracnose and Damping-off Diseases in Cucumber (Cucumis sativus) Seedlings. Ann. Appl. Biol. 2011, 159, 28–40. [Google Scholar] [CrossRef]
Figure 1.
Effect of sowing patterns on the severity of foot and root rot symptoms of pea (a) and wheat (b). n = number of assessed plants per treatment. Reported p-values resulted from the Kruskal–Wallis test. Root rot disease severity expressed as follows: for pea plants: healthy–asymptomatic plants (plants with a disease assessment of score 0), low (plants with scores of 1–2), moderate (plants with scores of 3–5), and high (plants with scores of 6–8) disease severity [12]; for wheat plants: no symptoms = plants with a score of 0, low = plants with lesions covering less than half of their stems, moderate = plants with lesions spanning from 50% to 100% of the stem circumference, and high = plants with a rotten/broken stem [26].
Figure 1.
Effect of sowing patterns on the severity of foot and root rot symptoms of pea (a) and wheat (b). n = number of assessed plants per treatment. Reported p-values resulted from the Kruskal–Wallis test. Root rot disease severity expressed as follows: for pea plants: healthy–asymptomatic plants (plants with a disease assessment of score 0), low (plants with scores of 1–2), moderate (plants with scores of 3–5), and high (plants with scores of 6–8) disease severity [12]; for wheat plants: no symptoms = plants with a score of 0, low = plants with lesions covering less than half of their stems, moderate = plants with lesions spanning from 50% to 100% of the stem circumference, and high = plants with a rotten/broken stem [26].

Figure 2.
The maximum likelihood phylogenetic tree (IQ-TREE) inferred from the partial β tubulin (tub2) gene sequence alignments used to verify the identity of D. pinodella isolates generated in this study (designated as FOEP and highlighted in red). Epitype and ex-type strains are marked with a superscript ‘T’. Branch support values determined via a Shimodaira–Hasegawa-like approximate likelihood ratio test (SH-aLRT) and ultrafast bootstraps (UFBoot) are shown above branches. The scale bar indicates 0.01 expected changes per site. The tree is rooted to Ascochyta pisi (CBS 122785).
Figure 2.
The maximum likelihood phylogenetic tree (IQ-TREE) inferred from the partial β tubulin (tub2) gene sequence alignments used to verify the identity of D. pinodella isolates generated in this study (designated as FOEP and highlighted in red). Epitype and ex-type strains are marked with a superscript ‘T’. Branch support values determined via a Shimodaira–Hasegawa-like approximate likelihood ratio test (SH-aLRT) and ultrafast bootstraps (UFBoot) are shown above branches. The scale bar indicates 0.01 expected changes per site. The tree is rooted to Ascochyta pisi (CBS 122785).

Figure 3.
The maximum likelihood phylogenetic tree (IQ-TREE) inferred from the partial TEF1 alpha gene sequence alignments used to verify identity of Fusarium isolates. The isolates generated in this study (designated as FOEP and highlighted in red) were accommodated with the Fusarium oxysporum species complex (FOSC) and the Fusarium tricinctum species complex (FTSC), which are highlighted in turquoise. Branch support values determined via a Shimodaira–Hasegawa-like approximate likelihood ratio test (SH-aLRT) and ultrafast bootstraps (UFBoot) are shown above branches. The scale bar indicates 0.08 expected changes per site. The tree is rooted to Neonectria ditissima (CBS 226.31).
Figure 3.
The maximum likelihood phylogenetic tree (IQ-TREE) inferred from the partial TEF1 alpha gene sequence alignments used to verify identity of Fusarium isolates. The isolates generated in this study (designated as FOEP and highlighted in red) were accommodated with the Fusarium oxysporum species complex (FOSC) and the Fusarium tricinctum species complex (FTSC), which are highlighted in turquoise. Branch support values determined via a Shimodaira–Hasegawa-like approximate likelihood ratio test (SH-aLRT) and ultrafast bootstraps (UFBoot) are shown above branches. The scale bar indicates 0.08 expected changes per site. The tree is rooted to Neonectria ditissima (CBS 226.31).

Figure 4.
Pre-emergence death and disease symptoms on 10 pea cultivars following infection with D. pinodella isolates. (a) Non-inoculated control plants; (b) pea plants inoculated with D. pinodella isolate FOEP 42.1500; (c) pea plants inoculated with D. pinodella isolate FOEP 42.1503; (d) pea plants inoculated with the five-isolate mixture; (e) rotten pea seeds recovered at harvest; (f) stunted plant and blackening of the transition zone tissue in pea cv. Baccara following inoculation with isolate FOEP 42.1503; (g) black necrotic lesions on tap roots of pea cv. Safran concentrated around the zone of seed attachment, completely encircling the tissue following inoculation with isolate FOEP 42.1501; (h) cross-section of infected pea stems; and (i) healthy root system in non-inoculated control plants (pea cv. Baccara). The order of the 10 pea cultivars in pictures (a–d), vertically in pots from left to right: Orchestra, Kagnotte, Safran, Poseidon, Kaplan, Furious, Kayanne, Isard, Casini, and Baccara.
Figure 4.
Pre-emergence death and disease symptoms on 10 pea cultivars following infection with D. pinodella isolates. (a) Non-inoculated control plants; (b) pea plants inoculated with D. pinodella isolate FOEP 42.1500; (c) pea plants inoculated with D. pinodella isolate FOEP 42.1503; (d) pea plants inoculated with the five-isolate mixture; (e) rotten pea seeds recovered at harvest; (f) stunted plant and blackening of the transition zone tissue in pea cv. Baccara following inoculation with isolate FOEP 42.1503; (g) black necrotic lesions on tap roots of pea cv. Safran concentrated around the zone of seed attachment, completely encircling the tissue following inoculation with isolate FOEP 42.1501; (h) cross-section of infected pea stems; and (i) healthy root system in non-inoculated control plants (pea cv. Baccara). The order of the 10 pea cultivars in pictures (a–d), vertically in pots from left to right: Orchestra, Kagnotte, Safran, Poseidon, Kaplan, Furious, Kayanne, Isard, Casini, and Baccara.

Figure 5.
(a) Effects of D. pinodella isolates on pea emergence and root rot disease severity. Data are presented across 10 pea cultivars. (b) Mean reaction per pea cultivar to isolates of D. pinodella. Root rot disease severity expressed as healthy–no symptoms (plants with a disease assessment score of 0), low (plants with scores of 1–2), moderate (plants with scores of 3–5), and high (plants with scores of 6–7) disease severity. Dead (disease assessment score of 8) = pre-emergence death was calculated relative to the corresponding non-inoculated control.
Figure 5.
(a) Effects of D. pinodella isolates on pea emergence and root rot disease severity. Data are presented across 10 pea cultivars. (b) Mean reaction per pea cultivar to isolates of D. pinodella. Root rot disease severity expressed as healthy–no symptoms (plants with a disease assessment score of 0), low (plants with scores of 1–2), moderate (plants with scores of 3–5), and high (plants with scores of 6–7) disease severity. Dead (disease assessment score of 8) = pre-emergence death was calculated relative to the corresponding non-inoculated control.

Figure 6.
Reactions of 10 pea cultivars to three D. pinodella isolates (FOEP 1500, 1501, and 1503) and the mixture of 5 isolates. Root rot disease severity expressed as healthy–no symptoms (plants with a disease assessment score of 0), low (plants with scores of 1–2), moderate (plants with scores of 3–5), and high (plants with scores of 6–7) disease severity. Dead (disease assessment score of 8) = pre-emergence death was calculated relative to the corresponding non-inoculated control.
Figure 6.
Reactions of 10 pea cultivars to three D. pinodella isolates (FOEP 1500, 1501, and 1503) and the mixture of 5 isolates. Root rot disease severity expressed as healthy–no symptoms (plants with a disease assessment score of 0), low (plants with scores of 1–2), moderate (plants with scores of 3–5), and high (plants with scores of 6–7) disease severity. Dead (disease assessment score of 8) = pre-emergence death was calculated relative to the corresponding non-inoculated control.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Pea cultivars used in this study.
| Cultivar Name | Cultivar Type 1 |
|---|---|
| Orchestra | Spring |
| Kagnotte | Spring |
| Poseidon | Spring |
| Kaplan | Spring |
| Kayanne | Spring |
| Baccara | Spring |
| Safran | Winter |
| Isard | Winter |
| Casini | Winter |
| Furious | Winter |
1 The pea cultivar descriptions are available at https://www.geves.fr/catalogue-france/; for the pea cv. ‘Baccara’, see Boutet et al., 2016 [44].
Table 2.
Isolation frequencies of predominant pathogens recovered from pea and wheat stems, crowns, and roots collected from the field experiment conducted in the 2018/2019 growing season at the experimental station of the National Research Institute for Agriculture, Food and Environment (INRAE)—Institute for Genetics, Environment, and Plant Protection (IGEPP) in Rennes, France.
Table 2.
Isolation frequencies of predominant pathogens recovered from pea and wheat stems, crowns, and roots collected from the field experiment conducted in the 2018/2019 growing season at the experimental station of the National Research Institute for Agriculture, Food and Environment (INRAE)—Institute for Genetics, Environment, and Plant Protection (IGEPP) in Rennes, France.
| Crop | Tissue | n 1 | D. pinodella | F. avenaceum | F. oxysporum | F. solani | F. equiseti |
|---|---|---|---|---|---|---|---|
| Pea | Stem | 54 | 85.2 a | 35.2 a | 3.7 b | 5.6 | 1.9 |
| Crown | 54 | 63.0 ab | 20.4 ab | 14.8 ab | 7.4 | 0.0 | |
| Root | 54 | 40.7 b | 13.0 b | 22.2 a | 11.1 | 3.7 | |
| Total no. of isolates | 177 | 102 | 37.0 | 22.0 | 13 | 3.0 | |
| % of plants affected | 92.6 | 40.7 | 35.2 | 22.2 | 5.6 | ||
| Wheat | Stem | 54 | 1.9 | 25.9 ab | 5.6 | 0.0 | 3.7 |
| Crown | 54 | 13.0 | 31.5 a | 7.4 | 1.9 | 9.3 | |
| Root | 54 | 1.9 | 9.3 b | 5.6 | 5.6 | 9.3 | |
| Total no. of isolates | 71 | 9 | 36 | 10 | 4 | 12 | |
| % of plants affected | 14.8 | 38.9 | 14.8 | 7.4 | 18.5 |
1 n = total number of different pea and wheat plant parts used for isolations. For each crop separately, significant differences among means within a column are indicated by a different letter (generalised linear models with a binomial distribution and logit link function at p < 0.05, followed by Sa idak-adjusted LSMeans post hoc test). D. pinodella = Didymella pinodella (syn. Phoma medicaginis var. pinodella, Peyronellaea pinodella); F stands for Fusarium, e.g., Fusarium avenaceum. The isolation frequencies for the species F. equiseti, F. crookwellense, F. dimerum, and D. pinodes are not presented in the table due to their low isolation rates.
Table 3.
Variations in isolation frequencies of the most common pathogens recovered from pea and wheat roots as affected by the sowing pattern.
Table 3.
Variations in isolation frequencies of the most common pathogens recovered from pea and wheat roots as affected by the sowing pattern.
| Crop | Sowing Pattern | n 1 | D. pinodella | F. avenaceum | F. oxysporum | F. solani | F. equiseti |
|---|---|---|---|---|---|---|---|
| Pea | Pure stand | 18 | 83.3 | 33.3 | 27.8 | 22.2 | 5.6 |
| Alternate rows | 18 | 100.0 | 55.6 | 38.9 | 16.7 | 11.1 | |
| Full mix | 18 | 94.4 | 33.3 | 38.9 | 27.8 | 0.0 | |
| p-value * | 0.6 | 0.6 | 0.6 | 0.8 | 0.2 | ||
| Wheat | Pure stand | 18 | 11.1 | 27.8 | 11.1 | 0.0 | 5.6 |
| Alternate rows | 18 | 16.7 | 55.6 | 27.8 | 16.7 | 33.3 | |
| Full mix | 18 | 16.7 | 33.3 | 5.6 | 5.6 | 16.7 | |
| p-value | 0.9 | 0.7 | 0.2 | 0.2 | 0.2 |
Didymella pinodella syn. Phoma pinodella, Phoma medicaginis var. pinodella, and Peyronellaea pinodella). F stands for Fusarium, e.g., Fusarium avenaceum. 1 n = total number of plants used for isolations. * p-values used to examine the sowing pattern effects originate from generalised linear models (bayesglm), with a binomial distribution and logit link function performed for each pathogen separately.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Šišić, A.; Baćanović-Šišić, J.; Gamba, F.M.; Finckh, M.R. Didymella pinodella: An Important Pea Root Rot Pathogen in France to Watch Out For? J. Fungi 2024, 10, 44. https://doi.org/10.3390/jof10010044
AMA Style
Šišić A, Baćanović-Šišić J, Gamba FM, Finckh MR. Didymella pinodella: An Important Pea Root Rot Pathogen in France to Watch Out For? Journal of Fungi. 2024; 10(1):44. https://doi.org/10.3390/jof10010044
Chicago/Turabian StyleŠišić, Adnan, Jelena Baćanović-Šišić, Fernanda M. Gamba, and Maria R. Finckh. 2024. "Didymella pinodella: An Important Pea Root Rot Pathogen in France to Watch Out For?" Journal of Fungi 10, no. 1: 44. https://doi.org/10.3390/jof10010044
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.