
The Effect of External Treatment of Arabidopsis thaliana with Plant-Derived Stilbene Compounds on Plant Resistance to Abiotic Stresses

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
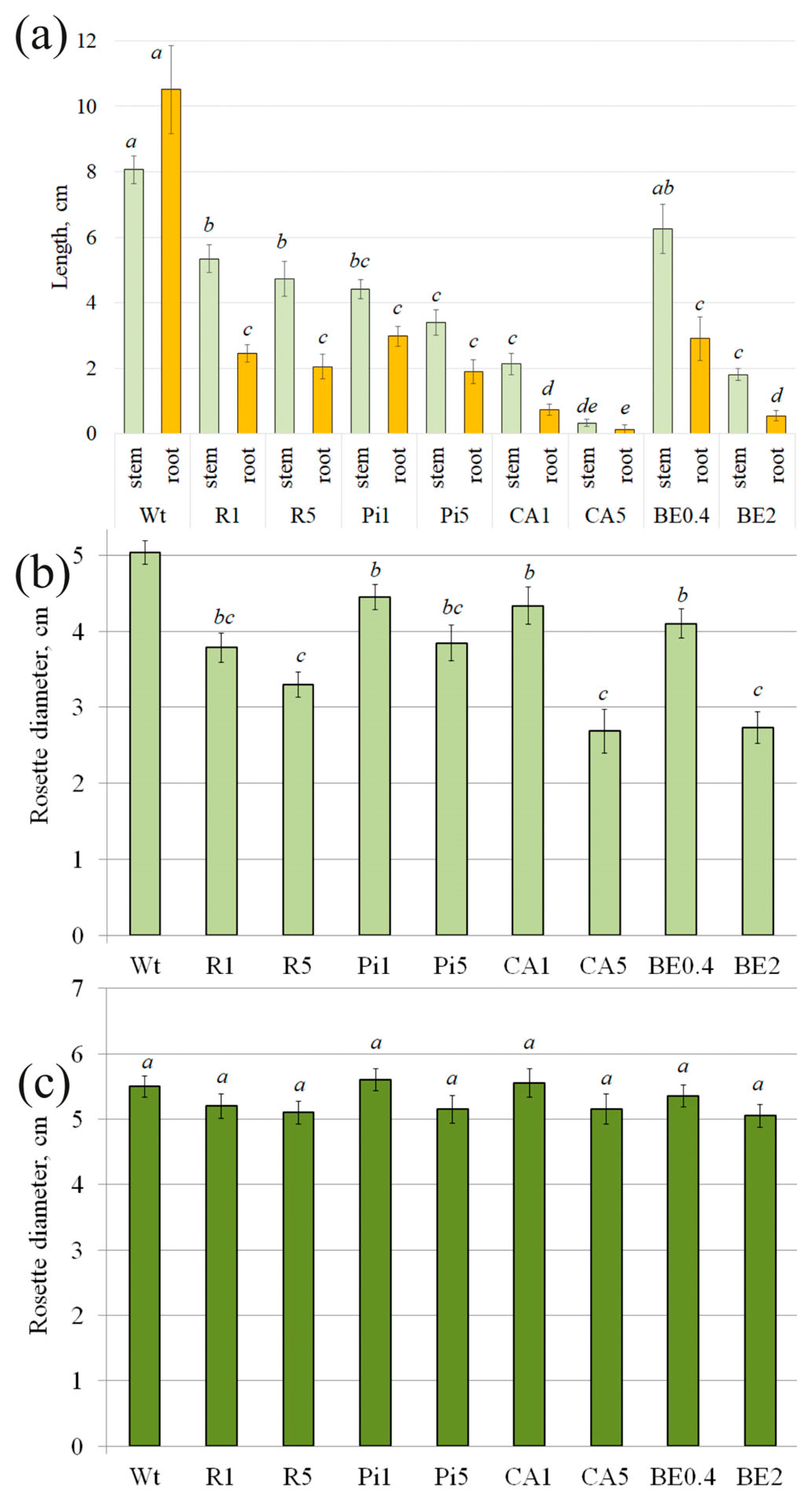
2.1. Effect of External Seed Treatment with Stilbenes and p-Coumaric Acid on A. thaliana Growth and Development
2.2. Influence of External A. thaliana Plant Treatment by Plant-Derived Stilbene Compounds on Survival Rate after Lethal Abiotic Stress Exposure
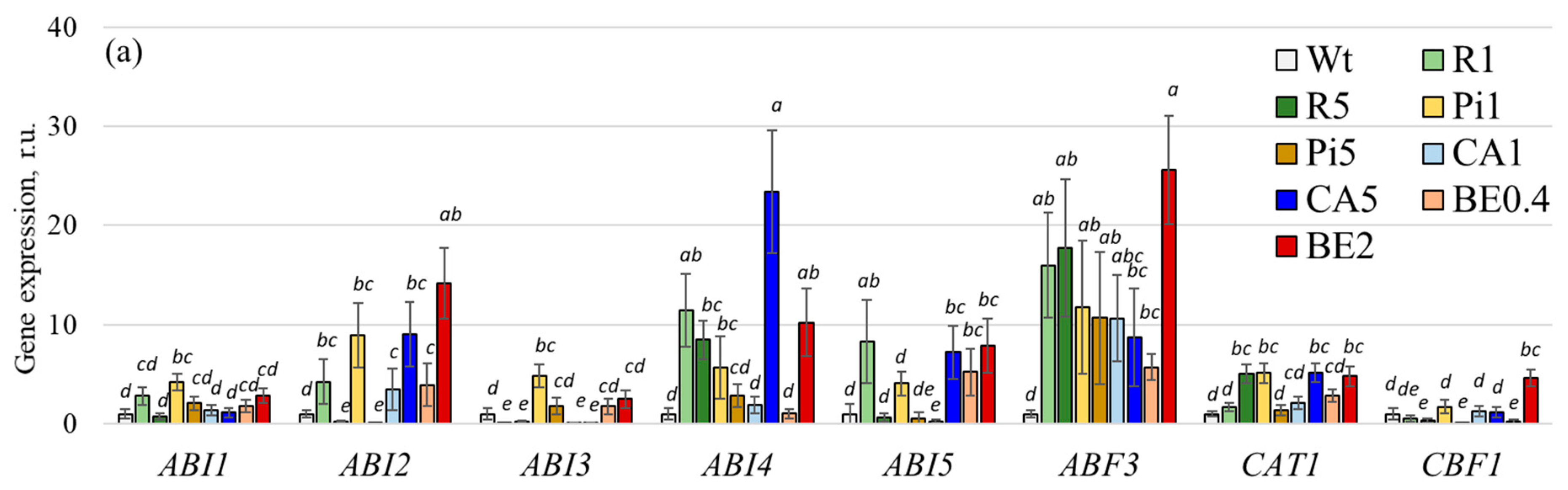
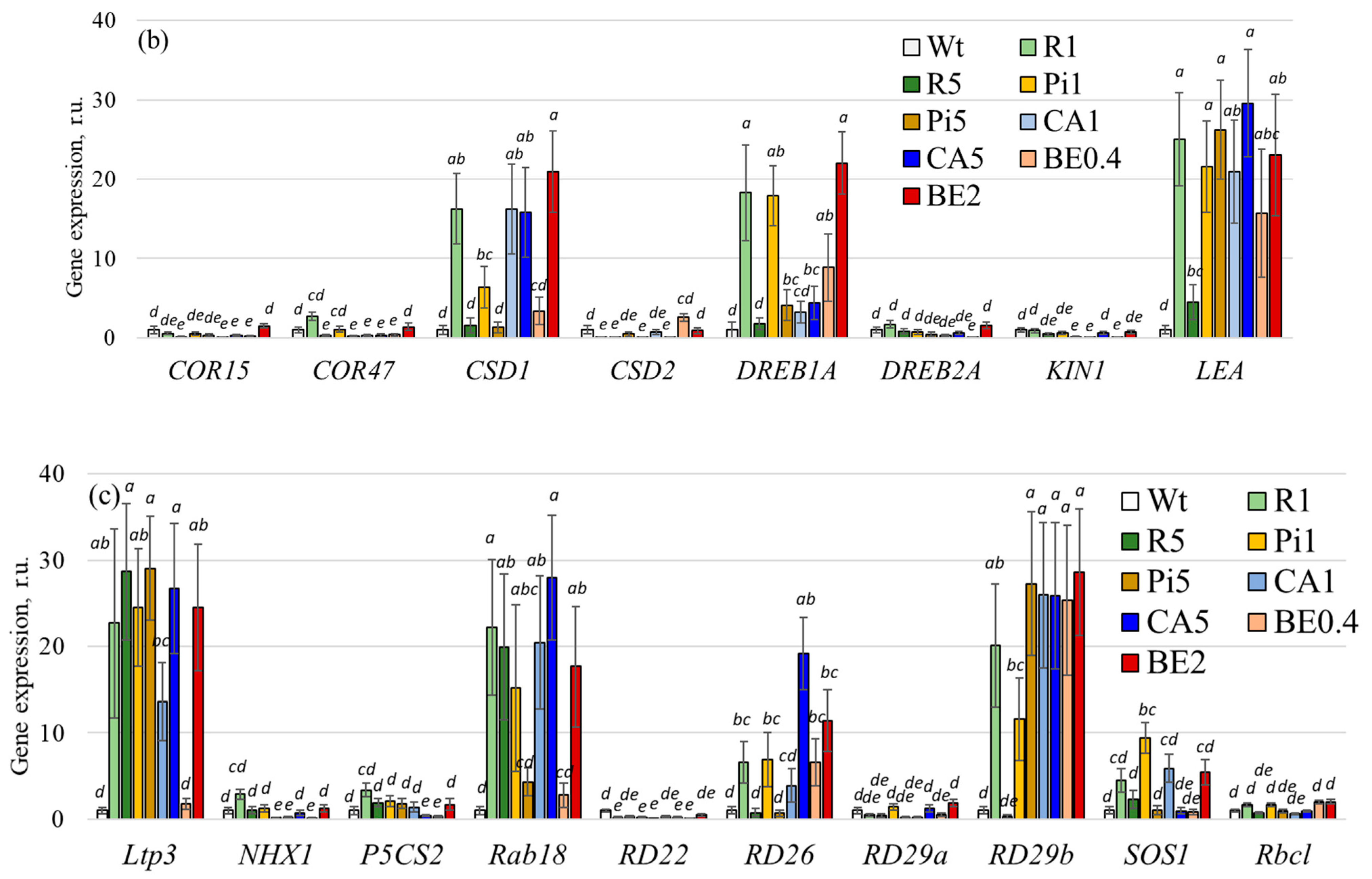
2.3. Alteration of Gene Expression of Phytohormone Metabolism and Stress-Associated Genes after Treatment with p-Coumaric Acid and Stilbene Compounds
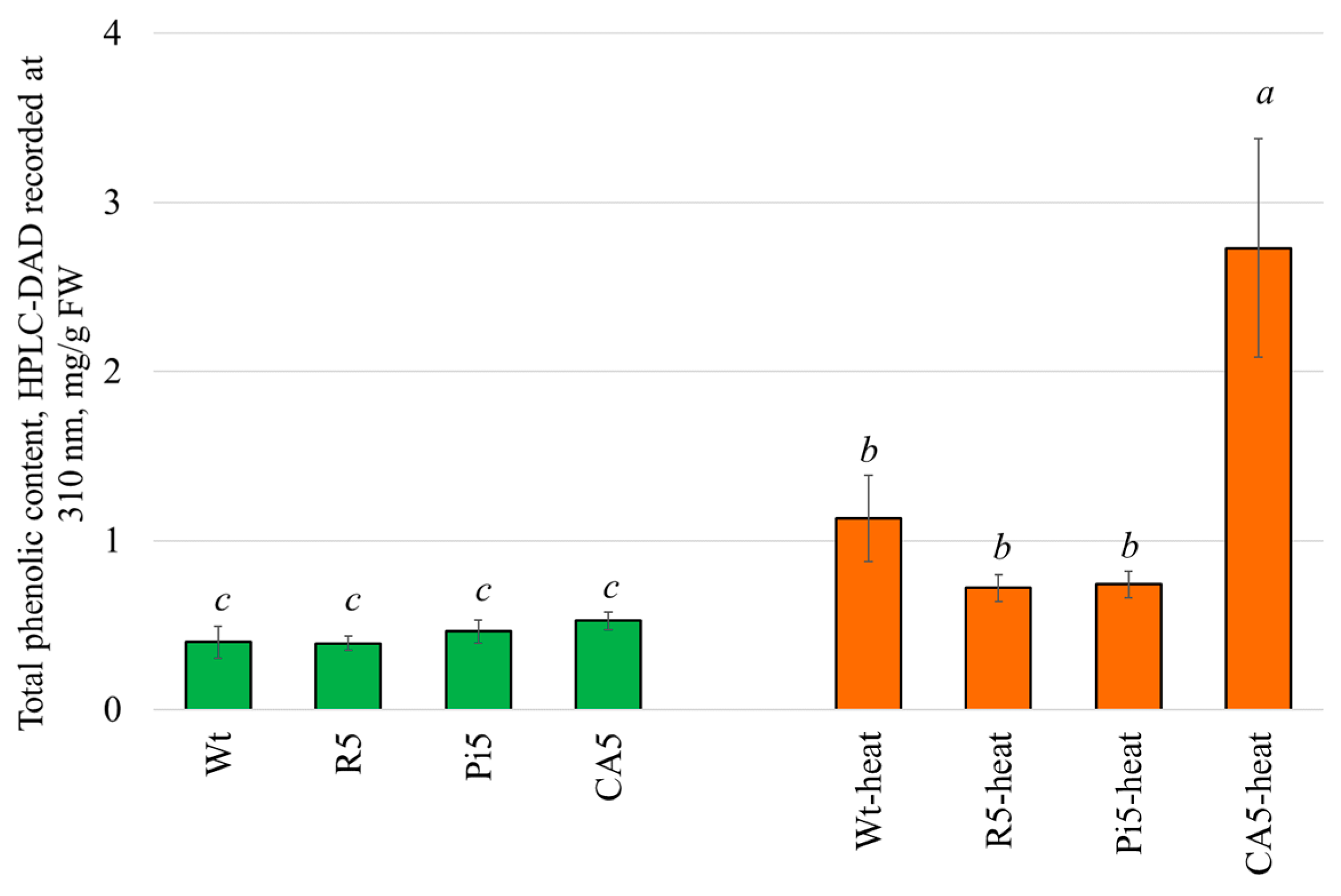
2.4. Changes in the Total Phenolic Content of the A. thaliana Plants after Heat, Stilbene, and p-Coumaric Acid Treatment
3. Conclusions
4. Materials and Methods
4.1. Plant Material
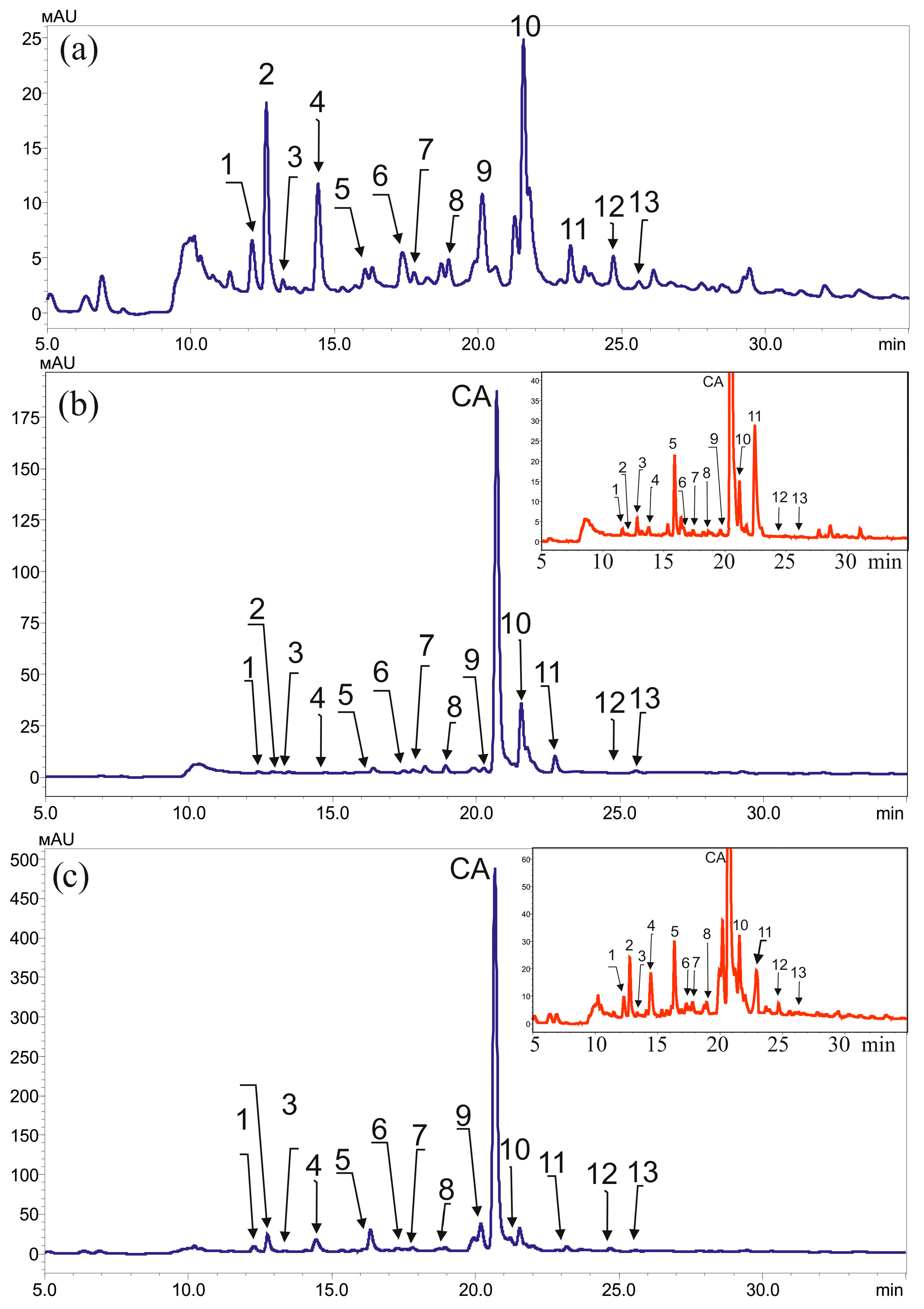
4.2. High-Performance Liquid Chromatography and Mass Spectrometry Analysis
4.3. RNA Extraction and Real-Time Quantitative PCR
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Al-Khayri, J.M.; Mascarenhas, R.; Harish, H.M.; Gowda, Y.; Lakshmaiah, V.V.; Nagella, P.; Al-Mssallem, M.Q.; Alessa, F.M.; Almaghasla, M.I.; Rezk, A.A. Stilbenes, a versatile class of natural metabolites for inflammation-an overview. Molecules 2023, 28, 3786. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V. Regulation of stilbene biosynthesis in plants. Planta 2017, 346, 597–623. [Google Scholar] [CrossRef]
- Valletta, A.; Iozia, L.M.; Leonelli, F. Impact of environmental factors on stilbene biosynthesis. Plants 2021, 10, 90. [Google Scholar] [CrossRef]
- Austin, M.B.; Bowman, M.E.; Ferrer, J.L.; Schröder, J.; Noel, J.P. An aldol switch discovered in stilbene synthases mediates cyclization specificity of type III polyketide synthases. Chem. Biol. 2004, 11, 1179–1194. [Google Scholar] [CrossRef]
- Schmidlin, L.; Poutaraud, A.; Claudel, P.; Mestre, P.; Prado, E.; Santos-Rosa, M.; Wiedemann-Merdinoglu, S.; Karst, F.; Merdinoglu, D.; Hugueney, P. A stress-inducible resveratrol O-methyltransferase involved in the biosynthesis of pterostilbene in grapevine. Plant Physiol. 2008, 148, 1630–1639. [Google Scholar] [CrossRef]
- Hall, D.; De Luca, V. Mesocarp localization of a bi-functional resveratrol/hydroxycinnamic acid glucosyltransferase of Concord grape (Vitis labrusca). Plant J. 2007, 49, 579–591. [Google Scholar] [CrossRef]
- Dry, I.B.; Robinson, S.P. Molecular cloning and characterisation of grape berry polyphenol oxidase. Plant Mol. Biol. 1994, 26, 495–502. [Google Scholar] [CrossRef]
- Rupprich, N.; Hildebrand, H.; Kindl, H. Substrate specificity in vivo and in vitro in the formation of stilbenes—Biosynthesis of rhaponticin. Arch. Biochem. Biophys. 1980, 200, 72–78. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.J. Cancer chemopreventive and therapeutic potential of resveratrol: Mechanistic perspectives. Cancer Lett. 2008, 269, 243–261. [Google Scholar] [CrossRef]
- Vingtdeux, V.; Dreses-Werringloer, U.; Zhao, H.; Davies, P.; Marambaud, P. Therapeutic potential of resveratrol in Alzheimer’s disease. BMC Neurosci. 2008, 9, S6. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering, and biological functions of stilbene phytoalexins in plants. Biofactors 2010, 36, 331–341. [Google Scholar] [CrossRef]
- Kiselev, K.V. Perspectives for production and application of resveratrol. Appl. Microbiol. Biotechnol. 2011, 90, 417–425. [Google Scholar] [CrossRef]
- Yin, W.; Shang, M.; Li, X.; Sang, S.; Chen, L.; Long, J.; Jiao, A.; Wang, J.; Jin, Z.; Qiu, C. Recent developments in sources, chemical constituents, health benefits and food applications of essential oils extracted from medicine food homology plants. Food Biosci. 2023, 55, 102997. [Google Scholar] [CrossRef]
- Gangopadhyay, A. Plant-derived natural coumarins with anticancer potentials: Future and challenges. J. Herb. Med. 2023, 42, 100797. [Google Scholar] [CrossRef]
- Pearce, R.B. Effects of exposure to high ozone concentrations on stilbenes in Sitka spruce (Picea sitchensis (Bong.) Carr.) bark and on its lignification response to infection with Heterobasidion annosum (Fr.) Bref. Physiol. Mol. Plant. Pathol. 1996, 48, 117–129. [Google Scholar] [CrossRef]
- Hovelstad, H.; Leirset, I.; Oyaas, K.; Fiksdahl, A. Screening analyses of pinosylvin stilbenes, resin acids and lignans in norwegian conifers. Molecules 2006, 11, 103–114. [Google Scholar] [CrossRef]
- Suprun, A.R.; Dubrovina, A.S.; Aleynova, O.A.; Kiselev, K.V. The bark of the spruce Picea jezoensis is a rich source of stilbenes. Metabolites 2021, 11, 714. [Google Scholar] [CrossRef]
- Shao, L.; Zhao, S.J.; Cui, T.B.; Liu, Z.Y.; Zhao, W. 2,3,5,4’- tetrahydroxystilbene-2-O-β-D-glycoside biosynthesis by suspension cells cultures of Polygonum multiflorum Thunb. and production enhancement by methyl jasmonate and salicylic acid. Molecules 2012, 17, 2240–2247. [Google Scholar] [CrossRef]
- Aleynova-Shumakova, O.A.; Dubrovina, A.S.; Manyakhin, A.Y.; Karetin, Y.A.; Kiselev, K.V. VaCPK20 gene overexpression significantly increased resveratrol content and expression of stilbene synthase genes in cell cultures of Vitis amurensis Rupr. Appl. Microbiol. Biotechnol. 2014, 98, 5541–5549. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Dubrovina, A.S.; Manyakhin, A.Y.; Karetin, Y.A.; Kiselev, K.V. Regulation of resveratrol production in Vitis amurensis cell cultures by calcium-dependent protein kinases. Appl. Biochem. Biotech. 2015, 175, 1460–1476. [Google Scholar] [CrossRef]
- Komaikul, J.; Kitisripanya, T.; Tanaka, H.; Sritularak, B.; Putalun, W. Enhanced Mulberroside A production from cell suspension and root cultures of Morus alba using elicitation. Nat. Prod. Commun. 2015, 10, 1253–1256. [Google Scholar] [CrossRef]
- Beekwilder, J.; Wolswinkel, R.; Jonker, H.; Hall, R.; de Vos, C.H.; Bovy, A. Production of resveratrol in recombinant microorganisms. Appl. Environ. Microbiol. 2006, 72, 5670–5672. [Google Scholar] [CrossRef]
- Dwibedi, V.; Kalia, S.; Saxena, S. Isolation and enhancement of resveratrol production in Xylaria psidii by exploring the phenomenon of epigenetics: Using DNA methyltransferases and histone deacetylase as epigenetic modifiers. Mol. Biol. Rep. 2019, 46, 4123–4137. [Google Scholar] [CrossRef]
- Galgano, F.; Caruso, M.; Perretti, G.; Favati, F. Authentication of Italian red wines on the basis of the polyphenols and bio-genic amines. Eur. Food Res. Technol. 2011, 232, 889–897. [Google Scholar] [CrossRef]
- Vitrac, X.; Monti, J.P.; Vercauteren, J.; Deffieux, G.; Merillon, J.M. Direct liquid chromatographic analysis of resveratrol de-rivatives and flavanonols in wines with absorbance and fluorescence detection. Anal. Chim. Acta 2002, 458, 103–110. [Google Scholar] [CrossRef]
- Suprun, A.R.; Dubrovina, A.S.; Tyunin, A.P.; Kiselev, K.V. Profile of stilbenes and other phenolics in Fanagoria white and red Russian wines. Metabolites 2021, 11, 231. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Ogneva, Z.V.; Aleynova, O.A.; Suprun, A.R.; Karetin, Y.A.; Dubrovina, A.S.; Kiselev, K.V. Tolerance of Arabidopsis thaliana plants overexpressing grapevine VaSTS1 or VaSTS7 genes to cold, heat, drought, salinity, and ultraviolet irradiation. Biol. Plant. 2021, 65, 111–117. [Google Scholar] [CrossRef]
- Lehmann, T.; Janowitz, T.; Sánchez-Parra, B.; Alonso, M.P.; Trompetter, I.; Piotrowski, M.; Pollmann, S. Arabidopsis NITRILASE 1 contributes to the regulation of root growth and development through modulation of auxin biosynthesis in seedlings. Front Plant Sci. 2017, 8, 36. [Google Scholar] [CrossRef]
- Sato, A.; Soeno, K.; Kikuchi, R.; Narukawa-Nara, M.; Yamazaki, C.; Kakei, Y.; Nakamura, A.; Shimada, Y. Indole-3-pyruvic acid regulates TAA1 activity, which plays a key role in coordinating the two steps of auxin biosynthesis. Proc. Natl. Acad. Sci. USA 2022, 119, e2203633119. [Google Scholar] [CrossRef]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin regulates the activity of reproductive meristems, flower organ size, ovule formation, and thus seed yield in Arabidopsis thaliana. Plant Cell. 2011, 23, 69–80. [Google Scholar] [CrossRef]
- Takei, K.; Yamaya, T.; Sakakibara, H. Arabidopsis CYP735A1 and CYP735A2 encode cytokinin hydroxylases that catalyze the biosynthesis of trans-Zeatin. J. Biol. Chem. 2004, 279, 41866–41872. [Google Scholar] [CrossRef]
- Wang, J.; Ma, X.M.; Kojima, M.; Sakakibara, H.; Hou, B.K. N-glucosyltransferase UGT76C2 is involved in cytokinin homeostasis and cytokinin response in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 2200–2213. [Google Scholar] [CrossRef] [PubMed]
- Curaba, J.; Moritz, T.; Blervaque, R.; Parcy, F.; Raz, V.; Herzog, M.; Vachon, G. AtGA3ox2, a key gene responsible for bioactive gibberellin biosynthesis, is regulated during embryogenesis by LEAFY COTYLEDON2 and FUSCA3 in Arabidopsis. Plant Physiol. 2004, 136, 3660–3669. [Google Scholar]
- Plackett, A.R.; Powers, S.J.; Fernandez-Garcia, N.; Urbanova, T.; Takebayashi, Y.; Seo, M.; Jikumaru, Y.; Benlloch, R.; Nilsson, O.; Ruiz-Rivero, O.; et al. Analysis of the developmental roles of the Arabidopsis gibberellin 20-oxidases demonstrates that GA20ox1, -2, and -3 are the dominant paralogs. Plant Cell. 2012, 24, 941–960. [Google Scholar] [CrossRef]
- Behnam, B.; Iuchi, S.; Fujita, M.; Fujita, Y.; Takasaki, H.; Osakabe, Y.; Yamaguchi-Shinozaki, K.; Kobayashi, M.; Shinozaki, K. Characterization of the promoter region of an Arabidopsis gene for 9-cis-epoxycarotenoid dioxygenase involved in dehydration-inducible transcription. DNA Res. 2013, 20, 315–324. [Google Scholar] [CrossRef]
- Milborrow, B.V. The pathway of biosynthesis of abscisic acid in vascular plants: A review of the present state of knowledge of ABA biosynthesis. J. Exp. Bot. 2001, 521, 1145–1164. [Google Scholar] [CrossRef]
- Barrero, J.M.; Rodríguez, P.L.; Quesada, V.; Piqueras, P.; Ponce, M.R.; Micol, J.L. Both abscisic acid (ABA)-dependent and ABA-independent pathways govern the induction of NCED3, AAO3 and ABA1 in response to salt stress. Plant Cell Environ. 2006, 29, 2000–2008. [Google Scholar] [CrossRef]
- Binder, B.M.; Mortimore, L.A.; Stepanova, A.N.; Ecker, J.R.; Bleecker, A.B. Short-term growth responses to ethylene in Arabidopsis seedlings are EIN3/EIL1 independent. Plant Physiol. 2004, 136, 2921–2927. [Google Scholar] [CrossRef] [PubMed]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [PubMed]
- Brocard-Gifford, I.; Lynch, T.J.; Garcia, M.E.; Malhotra, B.; Finkelstein, R.R. The Arabidopsis thaliana ABSCISIC ACID-INSENSITIVE8 encodes a novel protein mediating abscisic acid and sugar responses essential for growth. Plant Cell 2004, 16, 406–421. [Google Scholar] [CrossRef] [PubMed]
- Heidari, P. Comparative analysis of C-repeat binding factors (CBFs) in tomato and Arabidopsis. Braz. Arch. Biol. Technol. 2019, 62, e19180715. [Google Scholar] [CrossRef]
- Mizoi, J.; Kanazawa, N.; Kidokoro, S.; Takahashi, F.; Qin, F.; Morimoto, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arab. thaliana. J. Biol. Chem. 2019, 294, 902–917. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J. 2004, 39, 863–876. [Google Scholar] [CrossRef]
- Wang, Z.; Su, G.; Li, M.; Ke, Q.; Kim, S.Y.; Li, H.; Huang, J.; Xu, B.; Deng, X.P.; Kwak, S.S. Overexpressing Arabidopsis ABF3 increases tolerance to multiple abiotic stresses and reduces leaf size in alfalfa. Plant Physiol. Biochem. 2016, 109, 199–208. [Google Scholar] [CrossRef]
- Thomashow, M.F. Role of cold-responsive genes in plant freezing tolerance. Plant Physiol. 1998, 118, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Puhakainen, T.; Hess, M.W.; Mäkelä, P.; Svensson, J.; Heino, P.; Palva, E.T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Mol. Biol. 2004, 54, 743–753. [Google Scholar] [CrossRef]
- Mowla, S.B.; Cuypers, A.; Driscoll, S.P.; Kiddle, G.; Thomson, J.; Foyer, C.H.; Theodoulou, F.L. Yeast complementation reveals a role for an Arabidopsis thaliana late embryogenesis abundant (LEA)-like protein in oxidative stress tolerance. Plant J. 2006, 48, 743–756. [Google Scholar] [CrossRef]
- Fabro, G.; Kovács, I.; Pavet, V.; Szabados, L.; Alvarez, M.E. Proline accumulation and AtP5CS2 gene activation are induced by plant-pathogen incompatible interactions in Arabidopsis. Mol. Plant Microbe Interact. 2004, 17, 343–350. [Google Scholar] [CrossRef]
- Yang, H.; Postel, S.; Kemmerling, B.; Ludewig, U. Altered growth and improved resistance of Arabidopsis against Pseudomonas syringae by overexpression of the basic amino acid transporter AtCAT1. Plant Cell Environ. 2014, 37, 1404–1414. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, C.; Cheng, Y.; Ruan, M.; Ye, Q.; Wang, R.; Yao, Z.; Wan, H. Molecular evolution and functional divergence of stress-responsive Cu/Zn superoxide dismutases in plants. Int. J. Mol. Sci. 2022, 23, 7082. [Google Scholar] [CrossRef] [PubMed]
- Kurkela, S.; Borg-Franck, M. Structure and expression of Kin2, one of two cold- and ABA-induced genes of Arabidopsis thaliana. Plant Mol. Biol. 1992, 19, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Li, M.; Cheng, Z.; Xu, Q.; Song, X.; Shang, X.; Guo, W. Overexpression of a cotton nonspecific lipid transfer protein gene, GhLTP4, enhances drought tolerance by remodeling lipid profiles, regulating abscisic acid homeostasis and improving tricarboxylic acid cycle in cotton. Environ. Exp. Bot. 2022, 201, 104991. [Google Scholar] [CrossRef]
- Liu, H.; Tang, R.; Zhang, Y.; Wang, C.; Lv, Q.; Gao, X.; Li, W.; Zhang, H. AtNHX3 is a vacuolar K+/H+ antiporter required for low-potassium tolerance in Arab. thaliana. Plant Cell Environ. 2010, 33, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Zhao, X.; Fei, Y.; Zhang, X. Correlation of aquaporins and transmembrane solute transporters revealed by genome-wide analysis in developing maize leaf. Comp. Funct. Genom. 2012, 2012, 546930. [Google Scholar] [CrossRef]
- Harshavardhan, V.T.; Van Son, L.; Seiler, C.; Junker, A.; Weigelt-Fischer, K.; Klukas, C.; Altmann, T.; Sreebuvasyky, N.; Baumlein, H.; Kuhlmann, M. AtRD22 and AtUSPL1, members of the plant-specific BURP domain family involved in Arabidopsis thaliana drought tolerance. PLoS ONE 2014, 9, e110065. [Google Scholar] [CrossRef] [PubMed]
- Msanne, J.; Lin, J.; Stone, J.M.; Awada, T. Characterization of abiotic stress-responsive Arabidopsis thaliana RD29A and RD29B genes and evaluation of transgenes. Planta 2011, 234, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Nivison, H.; Stocking, C. Ribulose bisphosphate carboxylase synthesis in barley leaves. Plant Physiol. 1983, 73, 906–911. [Google Scholar] [CrossRef]
- Del Carmen Martínez-Ballesta, M.; Moreno, D.A.; Carvajal, M. The physiological importance of glucosinolates on plant response to abiotic stress in Brassica. Int. J. Mol. Sci. 2013, 14, 11607–11625. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr., mediates cold and drought stress tolerance. J. Plant Physiol. 2015, 185, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Aleynova, O.A.; Grigorchuk, V.P.; Dubrovina, A.S. Stilbene accumulation and expression of stilbene biosynthesis pathway genes in wild grapevine Vitis amurensis Rupr. Planta 2017, 245, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Ogneva, Z.V.; Volkonskaia, V.V.; Dubrovina, A.S.; Suprun, A.R.; Aleynova, O.A.; Kiselev, K.V. Exogenous stilbenes improved tolerance of Arabidopsis thaliana to a shock of ultraviolet B radiation. Plants 2021, 10, 1282. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Suprun, A.R.; Aleynova, O.A.; Ogneva, Z.V.; Kalachev, A.V.; Dubrovina, A.S. External dsRNA downregulates anthocyanin biosynthesis-related genes and affects anthocyanin accumulation in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 6749. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Suprun, A.R.; Ananev, A.A.; Nityagovsky, N.N.; Ogneva, Z.V.; Dubrovina, A.S.; Kiselev, K.V. Effect of calmodulin-like gene (CML) overexpression on stilbene biosynthesis in cell cultures of Vitis amurensis Rupr. Plants 2022, 11, 171. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Name | Wt | R5-C | P5-C | CA-C | Heat-C | Heat-R | Heat-P | Heat-CA |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Isorhanetin | 0 c | 0 c | 0.01 bc ± 0.01 | 0.01 bc ± 0.01 | 0.05 abc ± 0.02 | 0.02 bc ± 0.01 | 0.02 bc ± 0.01 | 0.10 a ± 0.04 |
| 2 | Indole-3-butyric acid | 0.02 c ± 0.01 | 0.01 c ± 0.01 | 0.01 c ± 0.01 | 0.01 c ± 0.01 | 0.16 ab ± 0.05 | 0.14 ab ± 0.04 | 0.10 b ± 0.02 | 0.25 a ± 0.08 |
| 3 | 7-Methysulfinylheptyl glucosinolate | 0.08 abc ± 0.03 | 0.04 bc ± 0.02 | 0.03 c ± 0.02 | 0.03 c ± 0.02 | 0.04 bc ± 0.01 | 0.03 c ± 0.01 | 0.04 bc ± 0.01 | 0.10 a ± 0.03 |
| 4 | 3-Indolylmethyl glucosinolate | 0.01 d ± 0.01 | 0.01 d ± 0.01 | 0.05 cd ± 0.01 | 0.01 d ± 0.01 | 0.21 ab ± 0.08 | 0.10 bc ± 0.02 | 0.06 cd ± 0.02 | 0.53 a ± 0.21 |
| 5 | Glucohirsutin | 0.01 c ± 0.01 | 0.06 bc ± 0.03 | 0.04 bc ± 0.01 | 0.07 b ± 0.03 | 0.06 b ± 0.02 | 0.04 bc ± 0.01 | 0.02 bc ± 0.01 | 0.65 a ± 0.15 |
| 6 | 4-Methoxy-3-indolylmethyl glucosinolate | 0.05 c ± 0.02 | 0.02 c ± 0.01 | 0.07 bc ± 0.02 | 0.03 c ± 0.02 | 0.10 ab ± 0.03 | 0.09 ab ± 0.03 | 0.09 ab ± 0.03 | 0.23 a ± 0.06 |
| 7 | Sinapoyl hexoside | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.04 a ± 0.02 |
| 8 | Kaempferol hexose dideoxyhexose | 0.01 c ± 0.01 | 0.02 bc ± 0.01 | 0.03 bc ± 0.01 | 0.08 ab ± 0.02 | 0.07 ab ± 0.02 | 0.03 bc ± 0.02 | 0.06 b ± 0.02 | 0.15 a ± 0.04 |
| 9 | Kaepferol-3-O-hexoside | 0.03 c ± 0.01 | 0.02 c ± 0.01 | 0.02 c ± 0.01 | 0.03 c ± 0.01 | 0.15 b ± 0.04 | 0.03 c ± 0.01 | 0.03 c ± 0.01 | 0.43 a ± 0.14 |
| 10 | Kaepferol-3,7-O-diramniside | 0.02 b ± 0.01 | 0.03 b ± 0.01 | 0.02 b ± 0.01 | 0.09 a ± 0.03 | 0.11 a ± 0.04 | 0.04 b ± 0.02 | 0.06 ab ± 0.03 | 0.08 ab ± 0.02 |
| 11 | Sinapoyl malate | 0.14 a ± 0.04 | 0.16 a ± 0.09 | 0.18 a ± 0.05 | 0.19 a ± 0.07 | 0.18 a ± 0.03 | 0.17 a ± 0.04 | 0.21 a ± 0.06 | 0.17 a ± 0.03 |
| 12 | 1,2-di-O-Sinapoyl-beta-O-glucose | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.04 a ± 0.01 | 0.01 a ± 0.01 | 0.02 a ± 0.01 | 0.01 a ± 0.01 | 0.03 a ± 0.01 | 0.02 a ± 0.01 |
| 13 | Sinapic acid | 0.04 a ± 0.02 | 0.02 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 | 0.01 a ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleynova, O.A.; Ogneva, Z.V.; Suprun, A.R.; Ananev, A.A.; Nityagovsky, N.N.; Beresh, A.A.; Dubrovina, A.S.; Kiselev, K.V. The Effect of External Treatment of Arabidopsis thaliana with Plant-Derived Stilbene Compounds on Plant Resistance to Abiotic Stresses. Plants 2024, 13, 184. https://doi.org/10.3390/plants13020184
Aleynova OA, Ogneva ZV, Suprun AR, Ananev AA, Nityagovsky NN, Beresh AA, Dubrovina AS, Kiselev KV. The Effect of External Treatment of Arabidopsis thaliana with Plant-Derived Stilbene Compounds on Plant Resistance to Abiotic Stresses. Plants. 2024; 13(2):184. https://doi.org/10.3390/plants13020184
Chicago/Turabian StyleAleynova, Olga A., Zlata V. Ogneva, Andrey R. Suprun, Alexey A. Ananev, Nikolay N. Nityagovsky, Alina A. Beresh, Alexandra S. Dubrovina, and Konstantin V. Kiselev. 2024. "The Effect of External Treatment of Arabidopsis thaliana with Plant-Derived Stilbene Compounds on Plant Resistance to Abiotic Stresses" Plants 13, no. 2: 184. https://doi.org/10.3390/plants13020184