
Surveillance of Landraces’ Seed Health in South Italy and New Evidence on Crop Diseases
by
 , and
, and
Eliana Dell’Olmo
1,
Massimo Zaccardelli
1,
Vincenzo Onofaro Sanaja
1,
Boris Basile
2 and
Loredana Sigillo
1,*
1
Council for Agricultural Research and Economics, Research Centre for Vegetable and Ornamental Crops, Via Cavalleggeri 25, 84098 Pontecagnano, Italy
2
Department of Agricultural Sciences, University of Naples Federico II, 80055 Portici, Italy
*
Author to whom correspondence should be addressed.
Plants 2023, 12(4), 812; https://doi.org/10.3390/plants12040812
Submission received: 31 December 2022
/
Revised: 7 February 2023
/
Accepted: 8 February 2023
/
Published: 11 February 2023
(This article belongs to the Section Plant Protection and Biotic Interactions)
Abstract
:During the last three years, more than 300 landraces belonging to different plant species have been the main focus of an Italian valorization research project (AgroBiodiversità Campana, ABC) aiming at analyzing, recovering, preserving, and collecting local biodiversity. In this context, phytosanitary investigation plays a key role in identifying potential threats to the preservation of healthy seeds in gene banks and the successful cultivation of landraces. The surveillance carried out in this study, in addition to highlighting the expected presence of common species-specific pathogens such as Ascochyta pisi in peas, Ascochyta fabae in broad beans, and Macrophomina phaseolina, Xanthomonas axonopodis pv. phaseoli, and Xanthomonas fuscans subsp. fuscans in beans, pointed to the presence of novel microorganisms never detected before in the seeds of some hosts (Apiospora arundinis in common beans or Sclerotinia sclerotiorum and Stemphylium vesicarium in broad beans). These novel seedborne pathogens were fully characterized by (i) studying their morphology, (ii) identifying them by molecular methods, and (iii) studying their impact on adult crop plants. For the first time, this study provides key information about three novel seedborne pathogens that can be used to correctly diagnose their presence in seed lots, helping prevent the outbreaks of new diseases in the field.
1. Introduction
Nowadays, most agricultural systems are based on the cultivation of commercial varieties that assure high mass yields, seriously limiting the preservation of biodiversity [1]. Much concern of the scientific community about ongoing biodiversity erosion derives from the observation that 70% of biodiversity has been lost during the last century and, even more alarmingly, most of the food consumed worldwide is produced from only 12 vegetable species [2,3,4,5]. This dramatic scenario has forced governments to act at a global and local scale to define strategies aiming to discover and preserve biodiversity [6,7]. With this aim, an increasing importance has been given recently to landraces as a tool to preserve agrobiodiversity. The landraces are defined as dynamic populations characterized by phenotypical diversity, genetic diversity, and adaptative features closely related to the environment where they have been developed [8,9]. Landraces represent an important resource because they are adapted to low-input agriculture and are suitable for cultivation in harsh environments [10]. The high specialization of landraces provides a pool of genes that can be used in plant breeding to increase the resistance of cultivated crops to abiotic stressors such as drought and floods, and to biotic stressors such as pests and diseases [3]. In addition, landraces play a key economic role in low-income countries, while in high-income nations they are considered as niche products [8,11], being perceived by the consumers as high-quality products [11]. Moreover, landraces were part of the cultural heritage of territories and were used in traditional food and local festivals [9]. Therefore, landraces are gaining importance due to their pivotal role in the cultural heritage of populations and their interesting properties, such as stability, adaptability, and nutritional values [4,8,9]. In order to preserve biodiversity and enhance knowledge about landraces, the global community has established guidelines such as the European Union Biodiversity Strategy to 2020 and the 2030 Agenda for Sustainable Development [6,12,13]. Accordingly, Italy established a national system for the conservation and enhancement of biodiversity with law n. 194 of 1 December 2015, which aimed at protecting local genetic resources from the risk of extinction and/or genetic erosion [6].
The increasing interest in landraces has led to the definition of international programs dedicated to their valorization and inclusion in germplasm banks [5], where plant material needs to be stored in perfect health conditions. One of the major issues linked to landrace conservation is the self-production of the seeds by local farmers, which often results in low-quality propagation material because of the presence of seedborne pathogens. Seeds are considered the first vehicle of pathogens in the environment and, in the worst case, they can introduce novel pathogens in a habitat or cause the re-emergence of diseases previously eradicated [14]. The successful storage of seeds in germplasm banks requires integrated seed health strategies that were developed all over the world [6] to mitigate the impact of seedborne pathogens on crops, and are based on the employment of secure propagation materials. In this context, the use of seeds that are certified disease-free or with a concentration of pathogens below imposed thresholds may represent a primary strategy to counteract seedborne diseases [15,16]. For this reason, there is an urgent need for the setup of methodologies aiming to provide diagnostic tools that are effective in detecting the complexity of pathogenic microflora. Knowledge of the biological cycles of the emerging pathogens and their aptitude for transmission from seeds to plants must also be sought to evaluate the real risk of epidemics [17].
The aim of the present work was to contribute to the knowledge about pathogenic microflora resident on seeds of Fabaceae, Brassicaceae, and Liliaceae landraces in order to provide new insights about the risks of losing biodiversity and the spread of seedborne disease outbreaks. Furthermore, this approach is oriented to (i) ensure the optimal phytosanitary conditions of seeds to be used as propagation materials, (ii) contribute to disease control measures, and (iii) limit the spread of pathogens in the environment. The research activity described in this work was carried out as part of the phytosanitary surveillance of the seed health of landraces that originated in the Campania region (Southern Italy) and are included on the priority list considered in the national project on biodiversity, “Agro Biodiversità Campana” (ABC), whose major aim was to analyze, recover, preserve, and collect local biodiversity. The results reported here complement those previously described for the fungi belonging to the Botryosphaeriaceae family associated with accessions of common bean (Phaseolus vulgaris L.) [18,19].
2. Results
2.1. Liliaceous Crops: Garlic (Allium sativum L.) and Onion (Allium cepa L.)
In this work, two accessions of garlic, “Bianco locale” and “Ecotipo locale di Caposele”(Table 1), were subjected to mycological screening on the cloves, which are the most commonly used propagation material for this crop. After seven days of plate incubation, fluffy, pink, Fusarium-like colonies developed from tissue fragments of “Bianco locale”. DNA extracted from a representative monoconidial culture (CREA OF 1119.1) was amplified in the ITS1 region. Using the primers listed in Table 2, the obtained sequence OQ148577 was aligned in GenBank, and the isolate was recognized as Fusarium proliferatum (Matsush. Nirenberg ex Gerlach & Nirenberg) after matching with the accession MK252904 of isolate PHVNO21 of F. proliferatum (Table 3).
Regarding three onion landraces (“Di Alife”, “Agostina”, and “di Eremiti”), screened for the presence of pathogenic fungi, the results showed that the only contamination was caused by the saprophytic Alternaria spp.. The BLASTn analyses performed with the ITS sequence identified the CREA OF 1115.1 (OQ280977) strain as A. alternata with an identity of 97.80% with strain SS_13 (MT497390) (Table 2).
2.2. Leguminous Crops and Novel Pathogen Reports
2.2.1. Cowpea (Vigna unguiculata (L.) Walp) and Common Bean (Phaseolus vulgaris L.)
The analyses carried out on V. unguiculata did not reveal the presence of fungal or bacterial pathogens. Conversely, all three accessions (“Corna dei Signori (San Marzano)”, “Fagiolino lungo” (San Marzano), and “Corna dei Signori” Caste S. Giorgio) resulted positive for a Bean Common Mosaic Virus (BCMV) infection after a Double Antibody Sandwich Enzyme Linked Immunosorbent Assay (DAS-ELISA) (see Supplementary Tables S1 and S2).
During the mycological screening on the P. vulgaris var. “Tondino di Castel di Sasso”, a white colony, not resembling known contaminants or key pathogens, developed on PDA plates. This strain, coded as CREA OF 1128.1, was selected for its ability to colonize the neighboring seeds and to inhibit their germination. The colony showed white and flat mycelium with black structures visible to the naked eye. Further analyses, performed with an optical microscope, highlighted the presence of globular-to-elliptic spores, brown or deep brown with an average diameter of 15 µm. Moreover, some agglomerates composed of conidiogenous cells were found, and they were arranged in variable shapes (Figure 1).
The ITS1 and β-tubulin partial sequences (OQ132873 and OQ249627) allowed us to identify the strain as Apiospora arundinis (Corda) Pintos & P. Alvarado. The sequences (Table 2) were aligned using the BLASTn tool in the GenBank database, and the results showed a 98.39% identity with A. arundinis isolate F4P4Fg (OM959567) in the ITS locus and an identity of 99.40% with the strain MUCL1684 (AB220322) for the β-tubulin sequence, confirming that the strain CREA OF 1128.1 is A. arundinis (homotopic synonym: Arthrinium arundinis) [33]. The pathogenicity tests validated Koch’s postulates, since the strain was able to cause disease in the common bean. Indeed, dried lesions on the foliage, ranging from creamy white to light brown, were observed 21 days after inoculation on 87% of the inoculated plants (Figure 2).
A phylogenetic tree was built with MEGA X by aligning the sequences of CREA OF 1128.1 for both ITS and β-tubulin sequences, with A. arundinis sequences retrieved from GenBank (Figure 3). The results confirmed that the CREA OF 1128.1 isolate belonged to the species A. arundinis, as the novel isolate grouped in clusters containing most of the A. arundinis strains for both ITS and TUB genes. In the ITS phylogenic tree, CREA OF 1128.1 grouped with Italian strains SF (MW361311), collected from marble, thus suggesting a putative common ancestor based on the geographic location. On the other hand, CREA OF 1128.1 showed similarity also with an Indian isolate from bamboo, thus disavowing the hypothesis of geographic location. Moreover, the ITS sequences formed more than one clade, indicating slight variability among the strains. A similar result was observed in the phylogenetic tree obtained by aligning the β-tubulin sequences. Indeed, the CREA OF 1128.1 strain fit in a group with 49% bootstrap with strains isolated in Switzerland and Germany, but also with isolates from China and Louisiana, and this result did not support the hypothesis that strains with common geographical origin were more closely related from a phylogenetic point of view. Moreover, in the β-tubulin region, genetic variability was flattened with respect to ITS. In both phylogenetic trees, the Diaporthe phaseolorum (Cooke & Ellis) Sacc. CMT 70 strain was used as an outgroup.
The sample of “Tondino di Castel di Sasso” also resulted infected by the isolate CREA_OF_ 1128.3 of Macrophomina phaseolina (Tassi) Goid. The colony appeared flat and black, and abundant microsclerotia were macroscopically visible. The mycelium presented hyphae placed at 90° and the connection among the main hypha and the branches was characterized by the typical constrictions near the septa. The identity was confirmed by DNA extraction and subsequent amplification with the specific primer pairs MpCalF-MpCalR and MpTefF-MpTefR [32], reported in Table 3, that generated bands of 525 and 400 bp, as expected. The results of the mycological screening are summarized in Table S1.
Regarding the thirty-seven P. vulgaris accessions screened for the presence of key pathogenic bacteria, it is worth reporting that in the landrace “Dente di Morto” the presence of Xanthomonas axonopodis pv. phaseoli (Xap) and X. fuscans subsp. fuscans (Xff) was revealed in eight accessions propagated in 2021, while the bacteria were not detected in 2020. Moreover, Xap was detected in the accession “dei Signori”. As a consequence of these results, the infected seed lots were retired from the collection.
Pseudomonas-type colonies were detected in the landrace “Rosso di Acerra”, but the in vivo tests revealed that the isolates could not be recognized as Pseudomonas savastanoi pv. syringae.
Finally, the DAS-ELISA tests showed the presence of (a) the viruses CMV in seven common bean accessions (“Cannellino Bianco di Calitri”, “Lardari”, and five accessions of “Dente di Morto”), (b) the Bean Common Mosaic Virus (BCMV) in seven accessions (four accessions of “della Regina”, “Dente di Morto Acc. 2020”, “Butirro Acc. 2021”, and “Tabaccanti”), and (c) of mixed infection (CMV/BCMV) in two accessions (di “Prata Melizzano” and “Rosso di Acerra”) (Supplementary Table S2).
2.2.2. Pea (Pisum sativum L.) and Lentils (Lens culinaris Medik.)
As expected, all eight pea accessions were found to be colonized by external contaminants such as Alternaria spp. (3% of contaminated seeds in one accession of “Centogiorni” and in one of “Santacroce”), Penicillium spp. and Cladosporium spp. (2% in “Santacroce Acc. T”), and Chaetomium spp. (44% in “Centogiorni Acc. 2021”). Ascochyta pisi Lib., a key pathogen, causing anthracnose in peas, was found in the pea accessions “Centogiorni from Pollenatrocchia” and “Santacroce accession M” (Table S1). The strains were coded as CREA OF 1122.4 and CREA OF 1122.5, respectively, and the sequence accession numbers obtained in GenBank were reported in Table 2. The BLASTn analyses conducted on the two strains revealed an identity of CREA OF 1122.4 (OQ148428) with the strain PA (MN653147) with a percentage of 99.16%, whereas the strain CREA OF 1122.5 (OQ148433) had an identity of 98.53% with the same GenBank accession, allowing us to identify the isolates as A. pisi.
No pathogenic fungi were detected from lentil seeds, whereas Fusarium-like colonies were isolated from diseased mother plants collected in the seed multiplication field (Table S1). The strain was named CREA OF 190.4 and identified as Fusarium oxysporum Schltdl., according to colony morphology and mycelium characteristics.
The isolate CREA OF 190.4 was analyzed by amplifying the ITS1 and TEF 1-α gene regions, using ITS1-4 and TEF688/1251 primer pairs. The PCR products were sequenced, and multiple alignments in GenBank were performed by using BLASTn. The isolates were finally identified as F. oxysporum f. sp. medicaginis, and the similarity was 100% for ITS (OQ134131) with the isolate XC02 (MT579855) and 100% with F. oxysporum f. sp. medicaginis isolate NRRL2254 (DQ837690.1) in the TEF 1-α locus (OQ249629) (Table 2).
The test performed to verify the pathogenicity of F. oxysporum CREA OF 190.4 revealed a slight virulence of the isolate on lentil local variety “di Colliano” (Figure 4). The symptoms obtained in laboratory experiments were represented by necroses at collar level with a size ranging from 2 to 5 mm. After this symptom appearance, the leaves initially turned yellow and finally dropped. In our experimental conditions, 56% of the inoculated plants showed the described symptoms, while the non-inoculated control remained healthy, suggesting a putative partial resistance in the accession “di Colliano”. Moreover, no disease development was observed in inoculated plants of the Medicago sativa variety “Legend”, contrasting the molecular identification of F. oxysporum f. sp. medicaginis.
Finally, the virological and bacteriological analyses showed that pea accessions were not infected by CMV or Pseudomonas syringae pv. pisi.
2.2.3. Broad Bean (Vicia faba L.) and Grass Pea (Lathyrus cicera L.)
In the case of the broad bean, in the accession “A Corna”, in addition to the common external contaminants, the key pathogen Ascochyta fabae Speg. (Teleomorph: Didymella fabae) was found, which is the causal agent of anthracnose [34] (Table S1). The identity of the strain, denominated CREA OF 1235.2, was confirmed by comparing ITS sequence OQ148403 with the nucleotide source in the NCBI database (a 96.16% identity was found with the isolate SA1, MN640757) (Table 2).
On the other hand, as a result of the mycological analyses of the broad bean local variety “Lunga (nocerino-sarnese) Acc. 2021”, the presence of a white colony with fluffy mycelium, able to grow on the seeds and inhibiting their germination, was observed in the isolation plates. Further morphological observation, revealing the presence of sclerotia, suggested that the colony CREA OF 1113.1 could be ascribed to Sclerotinia sclerotiorum (Lib.) de Bary, which is responsible for the white mold syndromes in several plants [35] (Table S1). The morphological analyses confirmed the colony features of Sclerotinia sp., showing (Figure 5) sclerotia with a mean diameter of 4.6 mm. Moreover, ascospores and apotecia with an average main diameter, respectively, of 9.0 µm and 5.2 mm were measured in the microscopic observation. The molecular analyses carried out using the ITS1-4 and Tef728-986 primer pairs confirmed the identification of CREA OF 1113.1 isolate as S. sclerotiorum. Indeed, BLASTn analyses showed that CREA OF 1113.1 isolate had a 99.09% identity with the isolate of S. sclerotiorum SCS18 (KY848721) in the ITS (OQ133464) region and 100% with the isolate ColPat-586 (MK089777) in the TEF 1-α sequence (OQ249628) (Table 2).
Since this is the first time that the pathogen was reported in the broad bean in Italy, pathogenicity tests were performed to confirm Koch’s postulates. The symptoms of the disease (leaf chlorosis and stem rot) were visible after one week from the inoculation. At the end of the experimental time (20 days post inoculation), 100% of broad bean plants showed chlorosis and necrosis on their foliage along with serious stem rot around and far from the inoculation point (Figure 6). In addition, the roots appeared necrotic (Figure 6).
The sequences obtained by the amplification of ITS, and TEF1-α regions by using the ITS1-4 and TEF728/986 primers pairs, were employed in a phylogenetic analysis making a comparison with sequences retrieved from the GenBank database, selected on the basis of plant host and geographic location. The strain Botrytis fabae EHF-Bf8 was used as a possible outgroup. The results obtained by analyzing the ITS region showed, as expected, that CREA OF 1113.1 formed a cluster with S. sclerotium, already reported in GenBank, with a significant bootstrap, indicating a limited genetic distance (Figure 7a). Similarly, the analyses of TEF1-α sequences (Figure 7b) showed that CREA OF 1128.1 grouped with 100% bootstrap support with accessions retrieved from GenBank obtained from different hosts, i.e., rosemary, rose, and soybean, originated in different geographic regions including the USA, Spain, and South America. Indeed, the observations suggested that, even if the sequences were selected based on different geographic locations and hosts, differences were not reflected in genetic variability.
During the mycological surveillance, Stemphylium vesicarium (Wallr.) E.G. Simmons [36] was isolated from the broad bean local variety “A Sciabola Acc. 2021” (Table S1). The colony was coded as CREA OF 1112.1 and characterized by a grey-brownish colony with slightly fluffy mycelium. Under microscopic observation, in our experimental condition, no conidia were developed. Molecular analyses were performed through the amplification of the gene region ITS by using the ITS1-4 and the TEF728-986 primer pairs. The obtained sequences were aligned in BLASTn, and the isolate was identified as S. vesicarium. Indeed, CREA OF 1112.1 showed an identity of 99.26% with S. vesicarium isolate KACC48911 (MN871526) in the ITS (OQ266773) and 99.20% for the TEF 1-α locus (OQ249630) with the isolate NA51 (MH628128). To confirm the pathogenicity, CREA OF 1112.1 isolate was subjected to in vivo assay on V. faba “A Sciabola Acc. 2021” and, seven days after the inoculation, the compatible reaction was evidenced as typical necrotic spots on 52% of the leaves, with concentric circles, that became larger during the incubation time (Figure 8).
From wild grass pea local variety “Maracuoccio”, a strain (CREA OF 355.4) of Stemphylium sp. was also isolated (Table S1). The isolate was identified molecularly as S. globuliferum (Vestergr.) E.G. Simmons [37] by amplifying and sequencing the TEF 1-α region (Table 2); after a BLASTn analysis, the CREA OF 355.4 (OQ148588) strain showed a 99.81% identity with the strain of S. globuliferum FMB-NSIA-TA(S) (MG020726). Pathogenicity tests were performed but no symptoms were obtained on tested plants; thus, it was considered not pathogenic on the species of isolation.
A phylogenetic tree was built to confirm the identity of S. vesicarium CREA OF 1112.1. The analyses were conducted by aligning the TEF 1-α sequence (Table 2) obtained for the isolates with others retrieved from the GenBank database, considering different hosts and geographical origins. The results confirmed the identity of CREA OF 1112.1 as S. vesicarium. Indeed, the isolates grouped with previously described strains in clusters with 54% bootstrap, probably due to the differences in the sequence length. Moreover, from the phylogenetic tree, no correlation between the strains based on geographic origin and hosts could be inferred (Figure 9).
The broad bean results were negative in bacteriological and virological analyses for the detection of P. syringae and CMV, suggesting that, for these local varieties, the main concern was represented by mycological seed infections. Similarly, pathogenic bacteria were not detected in the grass pea (Table S2).
2.3. Brassicaceae
Among the Brassicaceae accessions tested, the most interesting results were obtained in B. oleracea var. sabellica “Torzella riccia”, on which the mycological screening revealed the presence of several Alternaria spp. which represent key fungal pathogens for this species (Table S1). The molecular analyses of the ITS1 partial sequences helped to identify the diverse isolates. In detail, CREA OF 1231.1, CREA OF 1231.4, CREA OF 1231.5 and CREA OF 1231.6, respectively related to the sequence OQ148383, OQ148398, OQ148397 and OQ148399, were identified as Alternaria alternata (Fries) Keissler. Moreover, the ITS sequence of CREA OF 1231.2 (OQ302538) aligned with Alternaria infectoria E.G. Simmons [38] sequence MK562061 with 98.93% identity, and CREA OF 1231.3 was identified as Alternaria brassicicola Schwein, Indeed the sequence OQ134394 showed 100% identity with the accession MW386633. The accession numbers of the sequences uploaded in GenBank are summarized in Table 2.
In the same sample, the presence of Xanthomonas campestris pv. campestris (Xcc) (Pammel) Dowson was also evidenced (Table S2). The bacterial strains, coded as CREA OF 1231.20, CREA OF 1231.21, and CREA OF 1231.22, were isolated on mCS20ABN medium and resulted positive after PCR amplification according to the ISTA 7-019a protocol [23] using the DHL153/154 primer pair. Pathogenicity tests on B. oleracea variety “Palla di neve”, used as a positive control, confirmed the pathogenicity of the isolates, since the typical V-shaped necrotic lesions, surrounded by a chlorotic halo, were clearly observed on leaf veins (Figure 10).
3. Discussion
In the context of biodiversity preservation, this work evaluates health in seeds to be stored in germplasm banks and distributed to growers. The results of the screening carried out on Liliaceae, Fabaceace, and Brassicaceae accessions are discussed below.
To summarize, pathogenic fungi, bacteria, and viruses were present in 27%, 14%, and 27% of the accessions analyzed, respectively. In addition, novel pathogens were found in 3% of the examined accessions.
Regarding the Liliaceae crops, the analyses did not highlight pathogen contaminations in onion; while in garlic, F. proliferatum was detected. In Italy, this fungus was first reported, in the north, on garlic in 2011 [39] and subsequently was detected on the Welsh onion (Allium fistulosum) [40]. Therefore, the pathogen can be considered well established in Italy and is confirmed as a key pathogen for garlic. F. proliferatum is known to be one of the causal agents of Fusarium dry rot (FDR), a disease that asymptomatically affects Allium spp. in the field but that overall shows symptoms during postharvest. The asymptomatic behavior, associated with the vegetative propagation of the host, explains the dangerousness of the pathogen, which can survive for a long time in the clove tissues, compromising not only the storage but also the success of the new cultivations (where it is responsible for Fusarium basal rot) [41].
Regarding legumes, our study reported, for the first time in the common bean “Tondino di Castel di Sasso”, a pathogen identified as A. arundinis, which demonstrated its ability to infect the inoculated plants, causing necrotic spots on the leaves. The genus Apiospora is defined as cosmopolitan, being found in very different environments [42], and the reports of A. arundinis being a plant pathogen are very scarce. In Italy, it was found in the Florence Cathedral to cause the blackening of white marble [43] and it was isolated also from the environment in a library in Veneto [44]. The pathogenicity of this species was reported to cause different symptoms, such as a reddish stem in sugarcane and necrosis in bamboo in China, and necrotic lesions in rosemary in Iran [45,46,47]. This is the first report of A. arundinis on the common bean worldwide. It is speculated that the presence of this pathogen could be linked to climate change and to the temperature shift, which allows some microorganism to grow more easily with respect to previous decades. The same sample of “Tondino di Castel di Sasso” was contaminated by M. phaseolina, a necrotrophic seed-transmitted fungus, which caused charcoal rot and damping-off of the seedlings in the common bean and in over 500 other species [48]. In recent years, pathogen–pathogen interaction has been assumed to be the basis for the strengthening of disease expression, and several synergistic interactions are reported, also involving some other species of Botryosphaeraceae [49]. The control strategies against this pathogen are often frustrating because no resistance genes (inter alia, there is no applicable strategy in the case of landrace cultivation) or chemicals are available to counteract the disease. The persistence of microsclerotia as conservation organs and the polyphagous attitude of the pathogen nullify the effects of crop rotation. Conversely, an appropriate agronomical approach, mainly focused on the management of irrigation and favoring the preservation of beneficial microorganisms in the soil, can contribute to reduction of the disease incidence. It is clear that the use of certified seeds can limit the introduction of new inoculum in the fields.
As a result of the bacteriological analyses on P. vulgaris, the presence of X. fuscans subsp. fuscans (Xff) and X. axonopodis pv. phaseoli (Xap) was ascertained in the accession “dei Signori” and in most of the accessions of “Dente di morto”. Due to the high potential risk of transmission to plants and dissemination to the environment, the lots interested by the infection were removed from the collection. Moreover, diagnostic analyses were conducted on seed lots obtained in 2020 and the results indicated that Xap and Xff were not detected. This allowed for the recovery of healthy seed lots among those harvested in previous seasons. Lo Cantore et al. [50] reported that, already in 2002, Xap was found in southern Italy (Basilicata region) on the common bean landrace “Fagiolo di Sarconi”. In addition, in the same study the authors demonstrated a differential behavior of the plant materials tested against strains of Xap and Xff and concluded that the two landraces can be considered tolerant to the disease [50]. In 2014, the European Food Safety Authority (EFSA) determined that the risk associated with this pathogen ranged from moderate to high with several negative impacts on yield and quality [51]. Thereafter, the European and Mediterranean Plant Protection Organization (EPPO) highlighted its presence in all the European countries except for Finland, Sweden, and the United Kingdom [52], determining the shifting of the bacterium from quarantine pest to Regulated Non-Quarantine Pest (RNQP) [50].
Finally, BCMV was found in common bean and cowpea accessions [53]; in particular, on the former species, BCMV was sometimes associated with CMV. In Italy, mixed infection was documented in the Veneto region, starting from 2012 on the Lamon bean (P. vulgaris), an agricultural product obtained by the cultivation of four local varieties [54]. In addition, BCMV, as a single infective agent, was detected in 24% and 100% of the common bean and cowpea accessions tested, respectively. This high incidence in the cowpea is consistent with previous studies reporting that V. unguiculata is a major host of the virus [55]. In the case of viruses residing inside the embryo (such as CMV and BCMV), seed treatments with chemicals are often not effective and, in addition, resistance genes are present only in some genotypes. Several studies dealing with seed treatment with Plant Growth-Promoting Bacteria (PGPR) evidenced the PGPR’s effectiveness in improving the health of young plants and, consequently, their resistance to the early viral infections in V. unguiculata [56].
The analyses conducted on P. sativum revealed seed infection of A. pisi in the accessions “Centogiorni Acc. 2021” (from Pollenatrocchia) and “Santacroce Acc. M 2021”. A. pisi is one of the causal agents of anthracnose in the pea and it is one of the most studied pests in this species. The effect of seed infection was evaluated in Canada by Sivachandra et al. [57], who evidenced that the percentage of infection in seed lots (in the range 0.5–14.5%) was not related to seed germination or to the plant symptoms [57]. Conversely, the amount of rainfall after flowering seems to be associated with an increase in disease severity. Moreover, it was concluded that the transmission from the seeds to the plantlets did not significantly progress and that, in the case of A. pisi, the seeds represent mainly a way to store the pathogen, since it is present in all its parts [57]. The percentage of infection detected in this work on pea landraces is limited to 1%, thus indicating a phytosanitary status on this host that has to be monitored but that can be considered under control.
The seeds of lentil accessions analyzed in this study resulted noncontaminated by pathogenic fungi. Indeed, the major seedborne pathogens (Ascochyta lentils, Botrytis cinerea, and F. oxysporum f. sp. lentils) [25] resident on this species were not detected. Conversely, in the field located in Calitri (Salerno), pathogenic F. oxysporum was isolated from seed mother plants of local variety “di Colliano”. The isolates obtained in this work, after in vivo tests on a sample of “di Colliano” accession, caused slight crown rot and yellowing of leaves, but the molecular identification allowed us only to ascribe the strain to the F. oxysporum species. Usually, F. oxysporum f. sp. lentils is the causal agent of severe disease in lentils with symptoms of discoloration of the foliage and complete wilting of the plants [58]. This fungus was found in central Italy in 1999–2000 on a lentil local ecotype, but, according to the TEF sequencing done in our study, the strain found in Campania was identified as F. o. f. sp. medicaginis (Fome). Subsequently, this result was not confirmed by the inoculation on M. sativa variety “Legend”, whose susceptibility to Fusarium is unknown. On the other hand, the variety Yliki (Gaia seeds S.A., Greece), reported as susceptible to Fome [59], was not available for our research. Therefore, these results could be only considered as preliminary observations; further sequencing studies in different gene regions could provide more information about the taxonomy and the report of Fome in lentils.
The analysis of the phytosanitary status of broad bean seeds highlighted the presence of A. fabae, S. sclerotiorum, and S. vesicarium strains, and the pathogenicity tests carried out with these isolates demonstrated their ability to induce disease in plants. A. fabae (teleomorph Didymella fabae) is the causal agent of Ascochyta blight and was detected for the first time in 1991 in Cambridge (UK). According to the UK CAB International Distribution map [60], D. fabae was reported wherever the crop is cultivated (Italy included). The fungus attacks stems, leaves, fruit, and also the seeds, where it can be stored and transmitted to a new plant. Its spread in open fields can cause up to 95% yield losses in favorable conditions [61]. This makes an integrated disease control strategy, based on time of sowing and varietal choice, the most effective way to reduce the damage incidence in the crop [62]. Additionally, in this case, the use of pathogen-free seed is the first step for a successful cultivation. S. sclerotiorum is characterized by a wide host range, including several legumes in which it causes white mold, crown rot, and stem rot [35,63,64]. Moreover, S. sclerotiorum is known to produce severe economic and yield losses in the legume production systems [65,66], having a negative impact on seed quality [63]. In 2018, broad bean infection caused by S. sclerotiorum was reported in North Dakota with symptoms similar to those caused by the isolate CREA OF 1113.1 in laboratory tests, namely stem and root rot [67]. The CREA OF 1113.1 isolate was also involved in a phylogenetic analysis which confirmed the identity of the pathogen, since it grouped with strains previously isolated from different crops and geographical areas. The wide dissemination of this pathogen and the disease severity it causes suggest an urgent need for novel screenings that can identify and quickly counteract the infections [63]. In addition to S. sclerotiorum, in our study, strains of S. vesicarium were isolated from seeds of the broad bean and they showed the ability to produce the typical symptoms associated with Stemphylium disease. These observations are consistent with previous reports showing that Stemphylium spp. have a high variability in the host range, with some species characterized by a broad host range while others are very host-specific [37,68,69]. In this context, S. botryosum, S. solani and S. vesicarium, which are species commonly associated with legume diseases, were already reported in previous works [70,71,72,73,74,75,76]. Moreover, invasive Stemphylium infections on the broad bean were reported in 2016 in Australia [37], confirming the results obtained in this work. Our study represents the first report of the occurrence of S. sclerotiorum and S. vesicarium on the seeds of the broad bean in Italy.
Interestingly, following the mycological screening carried out during the monitoring, all the accessions of grass pea and wild grass pea (L. sativum and L. cicera) resulted noncolonized by fungal pathogens. The strain S. globuliferum CREA_OF_335.4 was isolated by wild grass pea (“Maracuoccio” landrace). Although the fungal species was previously recorded as a plant pathogen in M. sativa [77] and Trifolium alexandrinum L. [78], the strain isolated in this study resulted not pathogenic on the host in isolation. Thus, the “Maracuoccio” accession is judged as free from fungi that can compromise grass pea health, but a further characterization of CREA_OF_355.4 on alfalfa and clover could allow for the definition of its pathogenicity and the potential of its seeds as reservoir of the disease agent. The scientific literature on plant diseases in the grass pea is very scarce, probably due to its intrinsic resistance to plant pathogens. The Lathyrus spp. genome contains a pool of resistance genes able to counteract Ascochyta blight, Erysiphe spp., downy mildew (Peronospora lathyri-palustris), Cercospora pisi sativae, and Uromyces spp.. Moreover, wild Lathyrus species are also known to be resistant to broomrape (Orobanche crenata). According to a previous study, grass pea varieties can represent an alternative to traditional legume crops because of their agronomic traits and nutraceutical properties, and thanks to their resistance to biotic stress and drought and salinity tolerance [79].
The analyses conducted on Brassicaceae seeds highlighted that the most abundant isolates belonged to the genus Alternaria, including A. alternata and A. brassicicola; the latter is one of the commonly reported diseases on this plant species [80,81]. Moreover, the bacteriological analyses revealed the presence of Xanthomonas campestris pv. campestris (Xcc) on the local variety “Torzella riccia”. This is the first time that this bacterium has been reported on Brassica oleracea var. sabellica. Xcc represents one of the most common pathogens in the Brassicaceae family, including plants with high economic impact. It is endemic in central-southern Italy and it was diagnosed starting from the 1990s in broccoli landrace “Natalino”, in cauliflower and kohlrabi commercial varieties, and up to 2013 in the wild rocket [82,83]. As early as 1952, Xcc was reported as a bacterium primarily spread by seeds [84], and because of the ineffectiveness of chemicals and lack of resistance genes in local varieties, the importance of starting cultivation by using healthy propagation materials appears crucial [85].
In this work, the importance of seed health evaluation is evidenced. Many validated protocols are available to detect the key pathogens for which the seed is considered a pathway. No diagnostic procedures with minor economic impact are available for botanical species or for pathogens with limited diffusion, and for this reason, there is a risk that seed infection can escape phytosanitary control. When validated protocols were not available, the researchers had to balance the practical laboratory approach with the use of robust diagnostic procedures, within the limit of an appropriate budget, and taking into account the small number of seeds required for each accession to run the analyses. The ISF-regulated pest risk database, the EPPO Global Database https://www.seedtest.org/, accessed on 1 January 2020), and the ISTA Reference Pest List [17] represented powerful tools to plan the work. The use of molecular methods, such as ITS, TEF, and β-tubulin region sequencing, were confirmed to be effective to identify most of the isolates belonging to different fungal species, including pathogens reported for the first time on the hosts. In particular, in these cases, the molecular identification provided information to design the pathogenicity tests that are still considered the main tool to attribute the role of disease agent to a microorganism.
The seeds analyzed in this work showed the presence of common saprophytes that did not affect seed health, while key and novel pathogens were recorded in one third of the accessions. This confirmed the importance of full screening on seeds to be used as propagation materials, because it allows for the start of seed sanitation programs aiming to preserve and valorize the local varieties and their territories.
4. Materials and Methods
4.1. Sampling and Isolation of Fungal and Bacterial Strains from Seeds
Within the ABC project, landraces of Liliaceae (garlic and onion), Fabaceae (cowpea, common bean, pea, lentils, broad bean, and grass pea) and Brassicaceae (B. oleracea and B. rapa) were multiplied in the original area of cultivation (in situ) or in experimental farms (ex situ) with the aim of producing seeds to be stored in the germplasm bank. The list of the landraces taken into account in this study (in some cases landraces were represented by more than one accession), the type of analysis carried out on them, and the methods used are summarized in Table 1. The diagnostic studies were performed on 3 accessions of onion, 2 of garlic, 6 of Brassica spp., 5 of grass pea (representing two Lathyrus species), 37 of common bean (representing 27 landraces), 8 of pea (representing 2 landraces), 6 of broad bean (representing 3 landraces), and 3 of cowpea. For each species, the target pathogens (reported in Table 1) were chosen among the key pathogens listed in the ISF (International Seed Federation) regulated pest risk database [21] and for which the seed is considered a pathway. The pests never detected in Italian territory on the studied hosts, according to the EPPO Global Database, were excluded by the screening. For those vegetable species (garlic, grass pea, lentils, broad bean, and cowpea) not included in the ISF database, the criterion adopted to choose the target pests was based on the literature. For each accession, the phytosanitary status was evaluated on a representative sample of 300 seeds that was divided into three subsamples of 100 seeds, separately subjected to mycological, bacteriological, and virological analyses. In the case of garlic, whose commercial propagation material is represented by cloves, the sample consisted of 200 cloves, divided into 2 subsamples subjected to mycological and bacteriological analyses.
4.1.1. Mycological Screening
In the case of mycological examination, the seeds were previously externally disinfected by a mild treatment with NaClO (0.5% active chloride) for 5–10 min. This treatment allowed us to avoid the overgrowth of Rhizopus sp. during the incubation without significantly compromising the isolation of pathogenic fungi. Then, the seeds were screened for the presence of the target microorganisms (Table 1) by plating them on potato dextrose agar (PDA, BD S.p.a., Milan, Italy) amended with 100 μg/mL chloramphenicol, 50 μg/mL streptomycin, and 50 μg/mL neomycin (Sigma-Aldrich, Milan, Italy), and incubating them for 10 days at 25 °C. Although this protocol was used to detect specific pathogens, the plating of seeds on such generic medium allowed us to also isolate other fungal species (pathogenic or not). This mycological examination protocol will be subsequently referred to as “mycological screening”. The number of seeds incubated in the 90 mm Petri dishes varied depending on the botanical species studied: nine seeds of common bean, cowpea, pea, and grass pea, six seeds of broad bean, twelve seeds of lentils, and sixteen seeds of brassica or onion. For garlic, 100 cloves were surface-sterilized with NaClO at 1% active chloride for 15 min, washed three times in sterile distilled water, and left to dry under a sterile hood. Then, the basal part of each clove was cut, put onto PDA, and incubated for 10 days at 25 °C. During the plate incubation, fungal colonies developed and, before their overgrowth, the isolates were transferred to fresh PDA plates. Finally, the colonies were observed for their morphological features and also used for the molecular and phylogenetic analyses (Section 4.5) and for the pathogenicity tests (Section 4.6). The mycological screening was carried out on all the species and accessions.
4.1.2. Detection of Target Bacteria
The bacteriological analyses were carried out on garlic, common bean, broad bean, grass pea, cowpea, pea, and Brassica spp. seeds by adapting validated ISTA and ISHI-Veg protocols. In the case of the common bean and Brassica spp., the standard protocols ISTA 7-021, ISTA 7-023, and ISTA 7-019a (published in 2022) [23] were employed to detect Xap and Xff and Psp and Xcc, respectively. Some modifications were necessary to adapt the cited procedures because the seed samples available at the ABC gene bank were smaller than those recommended by ISTA protocols.
Briefly, 100 seeds were immersed in washing buffer (sterile saline plus Tween™ 20 (0.02% v/v) [23] and incubated at 4 °C overnight to allow the bacteria to be dispersed. Then, 1 mL of buffer was withdrawn and employed in a 10-fold dilution in the same buffer until dilution 10−3, as reported in the “General methods” of the ISTA protocols. Briefly, each dilution was prepared by pipetting 0.5 mL from seed extract into a tube containing 4.5 mL of sterile diluent, which was then mixed prior to the next dilution step. A new sterile pipette was used for each dilution step. Then, 100 µL of diluted suspension and of undiluted extract were plated out, in triplicate, on the following selective media: Milk tween (MT) and Modified Sucrose Peptone medium (MSP) for the isolation of Xap, Xff, P. s. pv. phaseolicola, and P. s. pv. syringae. After five days, Pseudomonas and Xanthomonas-like colonies were purified on generic medium (Nutrient Agar, NA) to be submitted to further steps of identification and characterization.
Similarly, a modified ISTA 7-019a protocol was adopted to detect Xcc in Brassica spp. on the semi-selective media mCS20ABN and FS [23]. One hundred seeds were suspended in 1 mL of pre-chilled (2–4 °C) sterile saline plus Tween™ 20 (0.02% v/v) and shaken for 2.5 h at room temperature at 125 rpm. Then, the suspension was diluted until 10−3 (as reported above) and 100 µL was plated out on isolation media. After five days, Xanthomonas-like colonies were purified on NA and submitted to identification and characterization analyses.
Furthermore, the semiselective media KBBCA (King’s B Boric Acid medium) and SNAC (Sucrose NaCl medium) were used to select P. syringae pv. pisi according to the modified ISHI-Veg protocol [86]. One hundred seeds (1 gr of seeds per 2.5 mL solution) were macerated in sterile saline (0.85% NaCl) for 18 h. After incubation, the samples were shaken, and two serial 10-fold dilutions were prepared as reported above. Then, 100 µL of dilutions and undiluted seed extract were plated out on the specific selective media. MSP and King B media were used to attempt isolation of putative Pseudomonas syringae pathogenic pathovars from broad bean, grass pea, and cowpea. Maceration was performed in sterile saline (0.85% NaCl, 2.5 mL per 1 g seeds) and carried out overnight at 4 °C. Serial dilutions were obtained as previously described, and putative Pseudomonas colonies were purified on NA medium to be subjected to characterization analyses.
Finally, no isolation was performed on garlic cloves for the search of Erwinia carotovora subsp. carotovora because no rotted tissues were observed in any accession.
4.2. Virological Analyses
The virological analyses were conducted by grow-out assay, i.e., growing seedlings in controlled conditions, observing putative symptoms, and performing DAS-ELISA assays [87]. The tests were executed from symptomatic leaves and from asymptomatic bulk samples. The bulk samples were obtained by collecting five pieces of leaf tissue from five different plants and grinding them together 1/10 w/v in extraction buffer (Polyvinylpyrrolidone (PVP) 20 g, ovalbumin 2 g, sodium sulphite anhydrous 1.3 g in 1 L of phosphate buffered saline (PBS), pH 7.2, plus 10% Tween 20), as reported in the EPPO Standard 7/125 [87]. Noninfected and infected leaf tissues represented the NPC (Negative Processing Control, the baseline OD value for negative samples) and PPC (Positive Processing Control) as described in “Best practice for ELISA assay in seed health test”, published by ISF [86]. Blank control (buffers used in ELISA) was employed to evaluate the background optical density value in the absence of tissues. The samples and the controls were examined in duplicate. The assays were performed in Nunc Immunoplate maxiSorp and the results were read by measuring the absorbance at 405 nm by using the BioRad Microplate Reader, Model 550 (Bio Rad, Hercules, CA, USA). A sample was considered positive if the mean of two wells was higher than the double of the NPC. If the absorbance ranged between the NPC value and its double, the test was repeated.
DAS-ELISA testing was applied to verify putative infections of AMV, CMV, BCMV, and BCMNV in common bean, CMV and BCMV in cowpea, and CMV in pea and broad bean by using Bioreba ELISA Reagent sets (AMV Reagent set 480, Article code (Art.) Nr.: 140565; CMV Reagent set 960, Art. Nr.: 160662; BCMV Reagent set 480, Art. Nr.: 162065, and BCMNV Reagent set 480, Art. Nr.: 162165) with lyophilized positive control included (AMV Positive control component (C.) 140553; CMV Positive control C. 160653; BCMV Positive control C. 162053; and BCMNV Positive control C. 162153), following the manufacturer instructions (BIOREBA AG, Reinach, Switzerland). Besides the positive controls, the commercial sets included IgG, the secondary antibody conjugate to alkaline phosphatase, and the microtiter plates reported above.
4.3. Isolation from Symptomatic Mother Plants: The Case of Lentils
In late spring 2020, the accession of lentils “di Colliano”, multiplied in two open fields in Calitri (Salerno, Italy), showed, with an incidence of 40%, symptoms of wilting on the aerial part and crown and root blackening. Symptomatic radical tissues from five plants were washed in tap water, externally disinfected with 5% NaClO for 20 min, washed again in sterile distilled water, and dried under a hood. Fragments of 2–3 mm were cut from disinfected tissues and placed in PDA amended with antibiotics, as reported in Section 4.1. The plates were finally incubated at 25 °C in the dark for seven days. Fusarium-type colonies were transferred to fresh PDA plates and subjected to pathogenicity testing and molecular identification (Section 4.5 and Section 4.6).
4.4. Morphological Characterization of Fungal Colonies
The morphological characterization of fungal colonies was carried out macroscopically by observing the mycelium color and shape and the time required to overgrow the plate, whereas microscopic analyses were performed with an optical microscope, the Eclipse Nikon 90i with 10×, 20×, or 40× magnifications. Saprophyte fungi were recognized at the genus level according to the morphology features described by Champion [88]. The novel pathogens were characterized by colony morphology and, depending on the species, by microscopic specific features [89,90] such as: conidia shape and dimensions, hyphal morphology, mycelial color, presence/absence of sclerotia, presence/absence of sporangia, conidiophores, and conidiogenous cells. The dimensions of the specific structures were reported as a mean of twenty measurements.
4.5. Molecular Analyses and Phylogenesis
The fungal colonies preliminarily identified as belonging to putative pathogenic species were characterized by molecular analyses. First, spores were suspended in sterile water and monosporic cultures were obtained by plating the 10-fold dilution on PDA plates, with the aim of distinguishing pure single colonies. When fungi did not produce spores, monohyphal colonies were obtained by transplanting single mycelium tips grown on minimal medium (water agar). Monohyphal and monosporic cultures were then transferred in Potato Dextrose Broth (PDB, BD S.p.a., Milan, Italy) for 72 h at 25 °C and, finally, the grown mycelia were filtered using a sterile gauze. The samples were frozen at −80 °C, lyophilized and subjected to DNA extraction using the Genomic DNA isolation kit (Norgen, Biotek Corp., Thorold, ON, Canada) and following the manufacturer instructions. The PCR amplification was performed using different primer pairs based on the expected species as reported in Table 3. The couple ITS1 (5′-TCCGTAGGTGAACCTGCG–-3′)–ITS4 (5′-TCCTCCGTCTATTGATATGC-3′) was used to amplify the Internal transcribed spacer 1 region (ITS) from all the isolates studied. The primer pairs EF1–688F–EF1-1251R (5′-CGGTCACTTGATCTACAAGTGC-3′)/5′-CCTCGAACTCACCAGGTACCG-3′) and EF1–728F–EF1-968R (5′-CATCGAGAAGTTCGAGAAGG-3′/5′-TACTTGAAGGAACCCTTACC-3′) were employed to amplify the Translation elongation factor 1α (TEF-1α) sequences from Fusarium species and from all the other genera, respectively. Finally, the T1 (5′- TCCGTAGGTGAACCTGCGG–3′) and β-tub-2B (5′–ACCCTCAGTGTAGTGACCCTTGGC–3′) primer pair was used to amplify β-tubulin sequences of Apiospora sp.. For the PCR amplification, the Phusion™ High-Fidelity DNA Polymerase (2 U/µL) was employed according to manufacturer instructions and the PCR protocols were adapted for each primer pair. The PCR products were purified using the GeneJET PCR purification Kit (Thermofisher, Foster City, CA, USA) and sequenced by the Sanger method (BMR genomics, Padova, Italy). Sequences were trimmed and manually edited using Chromas Lite; they were identified using a nucleotide Basic Local Alignment Search Tool (BLAST) and then deposited in GenBank with the accession numbers reported in Table 2. Finally, the ITS, TEF 1-α, or β-TUB sequences were aligned using the default settings of Muscles, and then trimming was applied. The obtained alignments were employed to build phylogenetic trees using the max likelihood method and the Tamura–Nei model with 1000 bootstrap replicates using MEGA X.
Molecular identification of putative Xap, Xff, and Xcc was performed by using species-specific primer pairs, according to ISTA protocols mentioned above.
4.6. Pathogenicity Tests
The pathogenicity tests were carried out to verify Koch’s postulate only in the case of novel pathogen reports. Several inoculation protocols based on the pathosystems were employed, and the tests were performed in three replicates of ten plants, each belonging to the same local variety in which the isolates were found. The seed sample used for the pathogenicity assay was previously tested to ascertain the absence of pathogens. Negative controls were represented by plants treated with sterile water or agar, in the same conditions as those inoculated. Regarding A. arundinis CREA OF 1128.1, 10-day-old colonies were rinsed with 10 mL sterile deionized water and incubated for 10 min, to allow the spores to be dispersed; then, the spores were filtered, counted, and diluted in water to obtain a final suspension of 106 spores/mL. The inoculation was performed in two replicates on ten common bean plants (“Tondino di Castel di Sasso” landrace), at the trifoliate stage, by spraying the suspension on both the lower and upper pages of the leaves to allow the inoculum to go through the stomata. The plants were incubated in RH up to 90% in plastic boxes in greenhouse conditions at 25 ± 2 °C, until symptoms were visible on the foliage. The non-inoculated control was obtained by spraying the leaves with distilled sterile water. The data were recorded as an average percentage of diseased plants in the two replications.
S. sclerotiorum CREA OF 1113.1 was inoculated on 3-week-old broad bean plants (local variety “Lunga”) by the V-shape method [91], which consists in cutting the plant stem and applying mycelium plugs of 10-day-old colonies on the wounds. Sterile mycelium was used as a negative control. Laboratory film was used to seal the inoculum and the experiments were conducted at 26 °C with a 12 h photoperiod in 90% humidity; the irrigations were performed manually when necessary. The disease consisted in leaf chlorosis visible after one week from the inoculation and developed into aerial wilting and final stem and leaf necrosis. After 20 days, the symptoms were recorded as black necrotic lesions on the stems and the leaves and root rotting. The number of plants showing the described symptoms was counted and reported as an average percentage in the two replicates.
In the case of Stemphylium isolates, 10-day-old colonies were rinsed with 10 mL sterile deionized water and incubated for 10 min, to allow the mycelium fragments to be dispersed; then, the obtained suspension was diluted to 106 propagules/mL. The inoculation was performed on 3-week-old broad bean plants (“a Sciabola” landrace) by spraying the leaves at both the lower and upper page, to allow the inoculum to enter the stomata. Finally, the leaves were stinged with a sterile needle to facilitate the pathogen penetration. The plants were incubated in RH up to 90% in plastic boxes, in growth chamber conditions. Symptoms appeared 10 days after the inoculation and were visible on the foliage as brown spots. The non-inoculated control was obtained by spraying the leaves with distilled sterile water. The data were recorded as the percentage of spotted leaves in total on each plant and expressed as the mean in the sample.
Fusarium spp. colonies were inoculated in three-week-old lentil plants (“di Colliano” landrace) by dipping the roots in a 106 conidia/mL suspension, obtained from a 10-day-old colony. Then, the plants were transplanted in a mixture of peat and sand (1:1 v/v) and grown in climatic chamber conditions at 26 °C with a 12 h photoperiod. Treatment with water was used as negative control, while the Medicago sativa variety “Legend” was used as a control to verify the pathogenicity of the strain that was molecularly identified as F. o. f. sp. medicaginis. The disease was recorded as the number of plants showing rot at the crown level with a consequent yellowing of the foliage; the data were expressed as an average percentage of diseased plants in the three replicates of six plants each.
The pathogenicity tests of bacterial isolates were carried out according to the ISTA7-023, 7-021 and 7-019a [23] protocols mentioned above. P. vulgaris, variety “Michelet”, was used as a susceptible control to establish the pathogenicity of Xanthomonas-like strains isolated from seed samples. According to the ISTA 7-021 protocol, the first trifoliate leaf was completely immersed in a 108 CFU/mL bacterial suspension for 5 min. Then, the inoculated plants were incubated at 25 °C in a phytotron for 10 days. The symptoms of the disease were registered as typical polygonal chlorotic leaf spots surrounding leaf vein necrosis. Sometimes, necrotic streaks were also visible on the stems of inoculated plants. Plants treated with water represented the non-inoculated control. The strain of Xanthomonas aonopodis pv. phaseoli. LMG7455 was used as positive reference control.
The common bean variety “Michelet” was also used to test the pathogenicity of putative Pseudomonas. savastanoi. pv. phaseolicola isolate. Bean seeds were sown on sterile wet filter paper for 5 days and, at the beginning of the germination phase, the cotyledons were pricked with a sterile toothpick immersed in a 24-h-pure colony. Ten days after inoculation, the pathogenicity was assessed by observing the wounds on cotyledons. In the case of dried necrotic lesions, the strain was considered non-pathogenic. On the contrary, when hydropic halos surrounded the wounds, the strains were judged to be pathogenic. The strain of P. s pv. phaseolicola NCPPB 52.0 was used as a reference pathogenic strain.
Brassica oleracea var. botrytis variety “Palla di neve” was used as a susceptible control in the Xcc pathogenicity assays. In this case, sterile toothpicks were immersed in 24–48 pure Xcc-type colonies and used to prick the major veins of cauliflower leaves, at a point near the leaf edges. The plants were examined for the appearance of typical progressive V-shaped, yellow/necrotic lesions, with blackened veins, after 7–10 days at 25 °C. The strain PVCT 62.4 was used as a reference positive control. The pathogenicity tests for bacterial isolates were performed on six plants in three replicates.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12040812/s1. Table S1: Results of mycological screening; Table S2: Overview of bacteriological and virological results.
Author Contributions
Conceptualization, E.D. and L.S.; methodology, E.D., V.O.S., and L.S.; software, E.D.; validation, E.D. and L.S.; investigation, E.D. and L.S.; resources, M.Z.; data curation, E.D.; writing—original draft preparation, E.D.; writing—review and editing, B.B., E.D., and L.S.; supervision, L.S.; project administration, M.Z.; funding acquisition, M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Campania Region in the framework of the Rural Development Program 2014–2020, Measure 10.2.1.
Data Availability Statement
All data included in the main text and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tait, J.; Morris, D. Sustainable development of agricultural systems: Competing objectives and critical limits. Futures 2000, 32, 247–260. [Google Scholar] [CrossRef]
- Villa, T.C.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- Azeez, M.A.; Adubi, A.O.; Durodola, F.A.; Azeez, M.A.; Adubi, A.O.; Durodola, F.A. Landraces and Crop Genetic Improvement. In Rediscovery of Landraces as a Resource for the Future; Grillo, O., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Montesano, V.; Negro, D.; Sarli, G.; Logozzo, G.; Zeuli, P.S. Landraces in Inland areas of the Basilicata region, Italy: Monitoring and perspectives for on farm conservation. Genet. Resour. Crop Evol. 2012, 59, 701–716. [Google Scholar] [CrossRef]
- Bertacchini, E. Regional legislation in Italy for the protection of local varieties. J. Agric. Environ. Int. Dev. 2009, 103, 51–63. [Google Scholar] [CrossRef]
- Giupponi, L.; Pedrali, D.; Leoni, V.; Rodari, A.; Giorgi, A. The Analysis of Italian Plant Agrobiodiversity Databases Reveals That Hilly and Sub-Mountain Areas Are Hotspots of Herbaceous Landraces. Diversity 2021, 13, 70. [Google Scholar] [CrossRef]
- Decision adopted by the conference of the parties to the convention on biological diversity at its tenth meeting X/2. The Strategic Plan for Biodiversity 2011–2020 and the Aichi Biodiversity Targets. 2010. Available online: https://www.cbd.int/decision/cop/?id=12268 (accessed on 27 October 2022).
- Gibson, R.W. A review of perceptual distinctiveness in landraces including an analysis of how its roles have been overlooked in plant breeding for low-input farming systems. Econ. Bot. 2009, 63, 242–255. [Google Scholar] [CrossRef]
- Piergiovanni, A.R.; Margiotta, B. On farm survival of apulian legume and cereal landraces in relation to land cover/land use changes. A case study. Ital. J. Agron. 2021, 16, 1–9. [Google Scholar] [CrossRef]
- Pícha, K.; Navrátil, J.; Švec, R. Preference to Local Food vs. Preference to “National” and Regional Food. J. Food Prod. Mark. 2018, 24, 125–145. [Google Scholar] [CrossRef]
- Corrado, G.; Rao, R. Towards the Genomic Basis of Local Adaptation in Landraces. Diversity 2017, 9, 51. [Google Scholar] [CrossRef]
- EU Commission. EUROPE 2020—A Strategy for Smart, Sustainable and Inclusive Growth; EU Commission: Brussels, Belgium, 2010. [Google Scholar]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Piergiovanni, A.R.; Lioi, L. Italian Common Bean Landraces: History, Genetic Diversity and Seed Quality. Diversity 2010, 2, 837–862. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Plant Genetic Resources for Food and Agriculture Synthetic Account; The Second Report on The State of the World’s; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture (SoWPGR-2)|Knowledge for Policy. Available online: https://knowledge4policy.ec.europa.eu/publication/second-report-state-world’s-plant-genetic-resources-food-agriculture-sowpgr-2_en (accessed on 25 January 2023).
- Denancé, N.; Grimault, V. Seed pathway for pest dissemination: The ISTA Reference Pest List, a bibliographic resource in non-vegetable crops. EPPO Bull. 2022, 52, 434–445. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Building on Gender, Agrobiodiversity and Local Knowledge | International Treaty on Plant Genetic Resources for Food and Agriculture. Available online: https://www.fao.org/plant-treaty/tools/toolbox-for-sustainable-use/details/en/c/1178627/ (accessed on 25 January 2023).
- Dell’Olmo, E.; Zaccardelli, M.; Basile, B.; Corrado, G.; Sigillo, L. Identification and Characterization of New Seedborne Pathogens in Phaseolus vulgaris Landraces of Southern Italy. Pathogens 2023, 12, 108. [Google Scholar] [CrossRef] [PubMed]
- ISF Regulated Pest List Initiative—International Seed Federation. Available online: https://worldseed.org/resources/isf-regulated-pest-list-database/isf-regulated-pest-list-initiative/ (accessed on 25 January 2023).
- Epitome: International Journal of Multidisciplinary Research. Available online: http://epitomejournals.com/ (accessed on 25 January 2023).
- Common Names of Plant Diseases. Available online: https://www.apsnet.org/edcenter/resources/commonnames/Pages/default.aspx (accessed on 25 January 2023).
- International Seed Testing Association (ISTA). Seed Health Methods—International Rules for Seed Testing. Available online: https://www.seedtest.org/en/international-rules-for-seed-testing/seed-health-methods-product-1054.html (accessed on 29 December 2022).
- Infantino, A. Malattie fungine delle leguminose minori. In LEGUMINOSE MINORI (CECE, LENTICCHIA, CICERCHIA, FAVA) MONOGRAFIA—A Cura di Massimo Zaccardelli; MiPAAF: Rome, Italy, 2008; 83p. [Google Scholar]
- Garkoti, A.; Kumar, S.; Lal, M.; Singh, V. Major diseases of lentil: Epidemiology and disease management—A Review. Agriways 2013, 1, 62–64. [Google Scholar]
- Harveson, R.M.; Pasche, J.S.; Porter, L.; Chen, W.; Burrows, M. Compendium of Pea Diseases and Pests; American Phytopathological Society: Saint Paul, MN, USA, 2021; 130p. [Google Scholar]
- Omomowo, O.I.; Babalola, O.O. Constraints and Prospects of Improving Cowpea Productivity to Ensure Food, Nutritional Security and Environmental Sustainability. Front. Plant Sci. 2021, 12, 2392. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J.L.; Alves, A. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–13. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. AMPLIFICATION AND DIRECT SEQUENCING OF FUNGAL RIBOSOMAL RNA GENES FOR PHYLOGENETICS. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Santos, K.M.; Lima, G.S.; Barros, A.P.O.; Machado, A.R.; Souza-Motta, C.M.; Correia, K.C.; Michereff, S.J. Novel specific primers for rapid identification of Macrophomina species. Eur. J. Plant Pathol. 2020, 156, 1213–1218. [Google Scholar] [CrossRef]
- Dyko, B.J.; Sutton, B.C. New and interesting dematiaceous Hyphomycetes from Florida. Mycotaxon 1979, 8, 119–124. [Google Scholar]
- Ondřej, M.; Huňady, I. Faba bean (Vicia faba L.) breeding for resistance to anthracnose (Ascochyta fabae Speg.) in the Czech Republic. Czech J. Genet. Plant Breed. 2007, 43, 61–68. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.H.J.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Simmons, E.G. Perfect states of Stemphylium. Mycologia 1969, 61, 1–26. [Google Scholar] [CrossRef]
- Vaghefi, N.; Thompson, S.M.; Kimber, R.B.E.; Thomas, G.J.; Kant, P.; Barbetti, M.J.; van Leur, J.A.G. Multi-locus phylogeny and pathogenicity of Stemphylium species associated with legumes in Australia. Mycol. Prog. 2020, 19, 381–396. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria themes and variations (22-26). Mycotaxon 1986, 25, 287–308. [Google Scholar]
- Tonti, S.; Prà, M.D.; Nipoti, P.; Prodi, A.; Alberti, I. First Report of Fusarium proliferatum Causing Rot of Stored Garlic Bulbs (Allium sativum L.) in Italy. J. Phytopathol. 2012, 160, 761–763. [Google Scholar] [CrossRef]
- Alberti, I.; Prodi, A.; Montanari, M.; Paglia, G.; Asioli, C.; Nipoti, P. First report of Fusarium proliferatum associated with Allium fistulosum L. in Italy. J. Plant Dis. Prot. 2018, 125, 231–233. [Google Scholar] [CrossRef]
- Gálvez, L.; Palmero, D. Fusarium Dry Rot of Garlic Bulbs Caused by Fusarium proliferatum: A Review. Horticulturae 2022, 8, 628. [Google Scholar] [CrossRef]
- Pintos, A.; Alvarado, P. Phylogenetic delimitation of Apiospora and Arthrinium. Fungal Syst. Evol. 2021, 7, 197. [Google Scholar] [CrossRef]
- Santo, A.P.; Cuzman, O.A.; Petrocchi, D.; Pinna, D.; Salvatici, T.; Perito, B. Black on white: Microbial growth darkens the external marble of florence cathedral. Appl. Sci. 2021, 11, 6163. [Google Scholar] [CrossRef]
- Micheluz, A.; Manente, S.; Prigione, V.; Tigini, V.; Varese, G.C.; Ravagnan, G. The effects of book disinfection to the airborne microbiological community in a library environment. Aerobiologia 2018, 34, 29–44. [Google Scholar] [CrossRef]
- Monkai, J.; Phookamsak, R.; Tennakoon, D.S.; Bhat, D.J.; Xu, S.; Li, Q.; Xu, J.; Mortimer, P.E.; Kumla, J.; Lumyong, S. Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand. Diversity 2022, 14, 918. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Ghobadi Anvar, F. First report of leaf spot caused by Arthrinium arundinis on rosemary in Iran. J. Plant Pathol. 2014, 96, S4.126. [Google Scholar] [CrossRef]
- Liao, J.; Jiang, W.; Wu, X.; He, J.; Li, H.; Wang, T.; Cheng, L.; Chen, W.; Mo, L. First Report of Apiospora Mold on Sugarcane in China Caused by Apiospora arundinis (Arthrinium arundinis). Plant Dis. 2022, 106, 1058. [Google Scholar] [CrossRef] [PubMed]
- Marquez, N.; Giachero, M.L.; Declerck, S.; Ducasse, D.A. Macrophomina phaseolina: General Characteristics of Pathogenicity and Methods of Control. Front. Plant Sci. 2021, 12, 634397. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Venturi, V. Synergisms between microbial pathogens in plant disease complexes: A growing trend. Front. Plant Sci. 2015, 6, 385. [Google Scholar] [CrossRef]
- Cantore, P.L.; Giorgio, A.; Campion, B.; Iacobellis, N.S. Development of integrated disease control measures for the valorisation of traditional crops in Southern Italy: The case study of Fagioli di Sarconi. In The Sustainability of Agro-Food and Natural Resource Systems in the Mediterranean Basin; Vastola, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 145–158. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the pest categorisation of Xanthomonas axonopodis pv. phaseoli and Xanthomonas fuscans subsp. fuscans. EFSA J. 2014, 12, 3856. [Google Scholar] [CrossRef]
- EPPO. Xanthomonas phaseoli pv. phaseoli (XANTPH) [World distribution]—EPPO Global Database. Available online: https://gd.eppo.int/taxon/XANTPH/distribution (accessed on 27 December 2022).
- Wang, Y.; Gaba, V.; Yang, J.; Palukaitis, P.; Gal-On, A. Characterization of Synergy Between Cucumber mosaic virus and Potyviruses in Cucurbit Hosts. Phytopathology 2002, 92, 51–58. [Google Scholar] [CrossRef]
- Tarquini, G.; Martini, M.; Maestri, S.; Firrao, G.; Ermacora, P. The Virome of ‘Lamon Bean’: Application of MinION Sequencing to Investigate the Virus Population Associated with Symptomatic Beans in the Lamon Area, Italy. Plants 2022, 11, 779. [Google Scholar] [CrossRef]
- Aishwarya, P.; Rangaswamy, K.T.; Basavaraju, S.; Achari, R.; Govin, K.; Rudraswamy, K.P.; Prameela, H.A. Evaluation of the Seed-borne Nature of Bean Common Mosaic Virus (BCMV) in Cowpea. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 239–245. [Google Scholar] [CrossRef]
- Shankar, A.C.U.; Nayaka, S.C.; Niranjan-Raj, S.; Kumar, H.B.; Reddy, M.S.; Niranjana, S.R.; Prakash, H.S. Rhizobacteria-mediated resistance against the blackeye cowpea mosaic strain of bean common mosaic virus in cowpea (Vigna unguiculata). Pest Manag. Sci. 2009, 65, 1059–1064. [Google Scholar] [CrossRef]
- Sivachandra Kumar, N.T.; Banniza, S. Assessment of the effect of seed infection with Ascochyta pisi on pea in western Canada. Front. Plant Sci. 2017, 8, 933. [Google Scholar] [CrossRef] [PubMed]
- Jiskani, A.M.; Samo, Y.; Soomro, M.A.; Leghari, Z.H.; Ghulam Nabi Gishkori, Z.; Hussain Bhutto, S.; Qayoom Majeedano, A. A destructive disease of lentil: Fusarium wilt of lentil. Plant Arch. 2021, 21, 2117–2127. [Google Scholar] [CrossRef]
- Antonopoulos, D.F.; Elena, K. Susceptibility of Greek alfalfa and clover cultivars to Fusarium oxysporum f. sp. medicaginis and potential methods of disease control. J. Plant Dis. Prot. 2008, 115, 162–166. [Google Scholar] [CrossRef]
- CABI. Distribution Maps of Plant Diseases. Available online: https://www.cabi.org/publishing-products/distribution-maps-of-plant-diseases/ (accessed on 25 January 2023).
- Omri Ben Youssef, N.; Kerdraon, L.; Mieuzet, L.; Halila, I.; Jammezi, N.; Mbazia, A.; Kharrat, M.; Le May, C. Population structure of the faba bean blight pathogen Ascochyta fabae (teleomorph, Didymella fabae) in Tunisia. Phytopathol. Mediterr. 2019, 58, 81–94. [Google Scholar] [CrossRef]
- Ahmed, S.; Abang, M.M.; Maalouf, F. Integrated management of Ascochyta blight (Didymella fabae) on faba bean under Mediterranean conditions. Crop Prot. 2016, 81, 65–69. [Google Scholar] [CrossRef]
- Antwi-Boasiako, A.; Wang, Y.; Dapaah, H.K.; Zhao, T. Mitigating against Sclerotinia Diseases in Legume Crops: A Comprehensive Review. Agronomy 2022, 12, 3140. [Google Scholar] [CrossRef]
- Njambere, E.N.; Peever, T.L.; Vandemark, G.; Chen, W. Genotypic variation and population structure of Sclerotinia trifoliorum infecting chickpea in California. Plant Pathol. 2014, 63, 994–1004. [Google Scholar] [CrossRef]
- Derbyshire, M.C.; Denton-Giles, M. The control of sclerotinia stem rot on oilseed rape (Brassica napus): Current practices and future opportunities. Plant Pathol. 2016, 65, 859–877. [Google Scholar] [CrossRef]
- Danielson, G.A.; Nelson, B.D.; Helms, T.C. Effect of Sclerotinia Stem Rot on Yield of Soybean Inoculated at Different Growth Stages. Plant Dis. 2004, 88, 297–300. [Google Scholar] [CrossRef]
- Chapara, V.; Chittem, K.; Mendoza, L.E.D.R. First Report of White Mold Caused by Sclerotinia sclerotiorum on Faba Beans in North Dakota. Plant Dis. 2018, 102, 1669. [Google Scholar] [CrossRef]
- Hanse, B.; Raaijmakers, E.E.M.; Schoone, A.H.L.; van Oorschot, P.M.S. Stemphylium sp., the cause of yellow leaf spot disease in sugar beet (Beta vulgaris L.) in the Netherlands. Eur. J. Plant Pathol. 2015, 142, 319–330. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Hanse, B.; van Leeuwen, G.C.M.; Groenewald, J.Z.; Crous, P.W. Stemphylium revisited. Stud. Mycol. 2017, 87, 77–103. [Google Scholar] [CrossRef] [PubMed]
- Pantidou, M.E. Fungus-Host Index for Greece; CABI: Wallingford, UK, 1973. [Google Scholar]
- El-Buni, A.M.; Rattan, S.S. Flora of Libya. Suppl. In Check List of Libyan fungi; Al Faateh University: Tripoli, Libya, 1981. [Google Scholar]
- Cook, P.R.; Dube, A.J. Host-Pathogen Index of Plant Diseases in South Australia; Field Crops Pathology Group South Australian Department of Agriculture: Adelaide, Australia, 1989. [Google Scholar]
- Mułenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A Preliminary Checklist of Micromycetes in Poland; CABI: Wallingford, UK, 2008. [Google Scholar]
- Sheikh, F.; Dehghani, H.; Aghajani, M.A. Screening faba bean (Vicia faba L.) genotypes for resistance to Stemphylium blight in Iran. Eur. J. Plant Pathol. 2015, 143, 677–689. [Google Scholar] [CrossRef]
- Foister, C.E. The economic plant diseases of Scotland. Tech. Bull. Dep. Agric. Scot. 1961, 1, 1–210. [Google Scholar]
- Orieux, L.; Felix, S. List of Plant Diseases in Mauritius; Phytopathological Papers; Commonwealth Mycological Institute: Kew, UK, 1968; Volume 7, 48p. [Google Scholar]
- Samac, D.A.; Willbur, J.; Behnken, L.; Brietenbach, F.; Blonde, G.; Halfman, B.; Jensen, B.; Sheaffer, C. First Report of Stemphylium globuliferum Causing Stemphylium Leaf Spot on Alfalfa (Medicago sativa) in the United States. Plant Dis. 2014, 98, 993. [Google Scholar] [CrossRef]
- Omar, A.F.; Hafez, Y.M.; Emeran, A.A. Occurrence of a New Leaf Spot Disease on the Egyptian Clover (Trifolium alexandrinum L.) Caused by Stemphylium globuliferum (Vestergren) in Egypt. Egypt. J. Biol. Pest Control 2015, 25, 89–96. [Google Scholar]
- Lambein, F.; Travella, S.; Kuo, Y.H.; Van Montagu, M.; Heijde, M. Grass pea (Lathyrus sativus L.): Orphan crop, nutraceutical or just plain food? Planta 2019, 250, 821–838. [Google Scholar] [CrossRef]
- EPPO. Alternaria brassicicola (ALTEBI) [Overview]—EPPO Global Database. Available online: https://gd.eppo.int/taxon/ALTEBI (accessed on 28 December 2022).
- Macioszek, V.K.; Gapińska, M.; Zmienko, A.; Sobczak, M.; Skoczowski, A.; Oliwa, J.; Kononowicz, A.K. Complexity of Brassica oleracea–Alternaria brassicicola Susceptible Interaction Reveals Downregulation of Photosynthesis at Ultrastructural, Transcriptional, and Physiological Levels. Cells 2020, 9, 2329. [Google Scholar] [CrossRef]
- Sigillo, L.; Esposito, S.; Tripodi, P.; Serratore, G.; Pane, C. Host range and molecular typing of Xanthomonas spp. strains isolated from wild rocket (Diplotaxis tenuifolia) in Italy. Eur. J. Plant Pathol. 2021, 160, 693–705. [Google Scholar] [CrossRef]
- Scortichini, M.; Rossi, M. Recurrent infections of Xanthomonas campestris pv. campestris on crucifers in Central-Southern Italy. Inf. Fitopatol. 1994, 12, 48–50. [Google Scholar]
- Cook, A.A.; Walker, J.C.; Larson, R. Studies on the disease cycle of black rot of crucifers. Phytopathology 1952, 42, 162–167. [Google Scholar]
- Frost, K.E.; Groves, R.L.; Charkowski, A.O. Integrated Control of Potato Pathogens Through Seed Potato Certification and Provision of Clean Seed Potatoes. Plant Dis. 2013, 97, 1268–1280. [Google Scholar] [CrossRef] [PubMed]
- ISF. Detection of Pseudomonas syringae pv. pisi on Pea Seed; Developed by ISHI-Veg; ISF: Nypn, Switzerland, 2020. [Google Scholar]
- EPPO. PM 7/125 (1) ELISA tests for viruses. EPPO Bull. 2015, 45, 445–449. [Google Scholar] [CrossRef]
- Champion, R. Identifier les Champignons Transmis par les Semences—RÉMI CHAMPION—Achat Livre ou Ebook | fnac. Available online: https://livre.fnac.com/a133884/REMI-CHAMPION-Identifier-les-champignons-transmis-par-les-semences (accessed on 19 December 2022).
- Crous, P.W.; Groenewald, J.Z. A phylogenetic re-evaluation of Arthrinium. IMA Fungus 2013, 4, 133–154. [Google Scholar] [CrossRef]
- Ordóñez-Valencia, C.; Ferrera-Cerrato, R.; Quintanar-Zúñiga, R.E.; Flores-Ortiz, C.M.; Guzmán, G.J.M.; Alarcón, A.; Larsen, J.; García-Barradas, O. Morphological development of sclerotia by Sclerotinia sclerotiorum: A view from light and scanning electron microscopy. Ann. Microbiol. 2015, 65, 765–770. [Google Scholar] [CrossRef]
- Ramírez, M.; Pérez, L.M.; Montealegre, J.R. Susceptibility of different grapevine (Vitis vinifera L.) cultivars to Diplodia seriata and Diplodia mutila. Int. J. Agric. Nat. Resour. 2018, 45, 93–98. [Google Scholar] [CrossRef]
Figure 1.
Morphology of A. arundinis CREA OF 1128.1: (a) colony on PDA; (b) details of black structures; (c) conidia of CREA OF 1128.1; (d) conidiogenous cells giving rise to conidia.
Figure 1.
Morphology of A. arundinis CREA OF 1128.1: (a) colony on PDA; (b) details of black structures; (c) conidia of CREA OF 1128.1; (d) conidiogenous cells giving rise to conidia.

Figure 2.
Common bean plants inoculated with A. arundinis CREA OF 1128.1: (a) non-inoculated control; (b,c) details of lesions on common bean leaves infected with A. arundinis CREA OF 1128.1.
Figure 2.
Common bean plants inoculated with A. arundinis CREA OF 1128.1: (a) non-inoculated control; (b,c) details of lesions on common bean leaves infected with A. arundinis CREA OF 1128.1.

Figure 3.
Phylogenetic analysis based on ITS (a) or β-tubulin (b) sequences of Apiospora spp. isolates. ITS1 and β-tubulin sequences of D. phaseolorum CMT70 from Brazil were used as an outgroup. The bootstrap tree inferred from 500 replicates is shown. The phylogenetic tree was inferred by using the maximum likelihood method and the Tamura–Nei model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). Red squares highlight the position of A. arundins CREA OF 1128.1. Numbers in red indicates the bootstraps for the nodes and the scale bar was reported in blue. Phylogenetic analyses were conducted in MEGA X.
Figure 3.
Phylogenetic analysis based on ITS (a) or β-tubulin (b) sequences of Apiospora spp. isolates. ITS1 and β-tubulin sequences of D. phaseolorum CMT70 from Brazil were used as an outgroup. The bootstrap tree inferred from 500 replicates is shown. The phylogenetic tree was inferred by using the maximum likelihood method and the Tamura–Nei model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown next to the branches. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). Red squares highlight the position of A. arundins CREA OF 1128.1. Numbers in red indicates the bootstraps for the nodes and the scale bar was reported in blue. Phylogenetic analyses were conducted in MEGA X.

Figure 4.
Symptoms caused by F. oxysporum CREA OF 190.4 on lentil seedlings. In the left panel the detail of foliage yellowing was reported and, on the right, the crown rot symptoms.
Figure 4.
Symptoms caused by F. oxysporum CREA OF 190.4 on lentil seedlings. In the left panel the detail of foliage yellowing was reported and, on the right, the crown rot symptoms.

Figure 5.
S. sclerotiorum CREA OF 1113.1 morphological features: (a) colony morphology on PDA and sclerotia after 15 days of growth; (b) ascopores and apothecium; (c) sclerotium development.
Figure 5.
S. sclerotiorum CREA OF 1113.1 morphological features: (a) colony morphology on PDA and sclerotia after 15 days of growth; (b) ascopores and apothecium; (c) sclerotium development.

Figure 6.
Symptoms on broad bean plants inoculated with S. sclerotiorum CREA OF 1113.1 twenty days post inoculation. (a) Chlorotic leaves; (b) stem rot spread from the inoculation point; (c) necrosis and desiccation; (d) detailed view of stem rot; (e) details of root rot.
Figure 6.
Symptoms on broad bean plants inoculated with S. sclerotiorum CREA OF 1113.1 twenty days post inoculation. (a) Chlorotic leaves; (b) stem rot spread from the inoculation point; (c) necrosis and desiccation; (d) detailed view of stem rot; (e) details of root rot.

Figure 7.
Dendrogram illustrating the evolutionary history of the isolated CREA OF 1113.1 inferred by ITS (a) and TEF1-α (b) DNA sequencing. For each taxon, the graph reports the GenBank accession number, the three-letter code pathogen species, the two-letter ISO country code, and the host plant species within vertical lines. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances. Maximum Composite Likelihood method used to infer the phylogenetic tree. Red squares highlight the position of S. sclerotiorum CREA OF 1113.1. Numbers in red indicates the bootstraps for the nodes and the scale bar was reported in blue. Phylogenetic analyses were conducted in MEGA X.
Figure 7.
Dendrogram illustrating the evolutionary history of the isolated CREA OF 1113.1 inferred by ITS (a) and TEF1-α (b) DNA sequencing. For each taxon, the graph reports the GenBank accession number, the three-letter code pathogen species, the two-letter ISO country code, and the host plant species within vertical lines. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (500 replicates) is shown. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances. Maximum Composite Likelihood method used to infer the phylogenetic tree. Red squares highlight the position of S. sclerotiorum CREA OF 1113.1. Numbers in red indicates the bootstraps for the nodes and the scale bar was reported in blue. Phylogenetic analyses were conducted in MEGA X.

Figure 8.
S. vesicarium CREA OF 1112.1 symptoms on inoculated broad bean “A sciabola” local variety: (a) colony morphology of CREA OF 1112.1 on PDA; (b) necrotic spots on broad bean leaves.
Figure 8.
S. vesicarium CREA OF 1112.1 symptoms on inoculated broad bean “A sciabola” local variety: (a) colony morphology of CREA OF 1112.1 on PDA; (b) necrotic spots on broad bean leaves.

Figure 9.
The evolutionary history inferred for S. vesicarium CREA OF 1112.1 based on TEF 1-α was performed by using the maximum likelihood method and the Tamura–Nei model. The tree with the highest log likelihood (−665.65) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying the Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Tamura–Nei model, and then selecting the topology with the superior log likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). This analysis involved 13 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. There was a total of 733 positions in the final dataset. Red squares highlight the position of S. vesicarium CREA OF 1112.1. Numbers in red indicates the bootstraps for the nodes and the scale bar was reported in blue. Phylogenetic analyses were conducted in MEGA X.
Figure 9.
The evolutionary history inferred for S. vesicarium CREA OF 1112.1 based on TEF 1-α was performed by using the maximum likelihood method and the Tamura–Nei model. The tree with the highest log likelihood (−665.65) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying the Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Tamura–Nei model, and then selecting the topology with the superior log likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches). This analysis involved 13 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. There was a total of 733 positions in the final dataset. Red squares highlight the position of S. vesicarium CREA OF 1112.1. Numbers in red indicates the bootstraps for the nodes and the scale bar was reported in blue. Phylogenetic analyses were conducted in MEGA X.

Figure 10.
Infection of Xcc CREA OF 1231.20 in B. oleracea var. botrytis; (a) non-inoculate control; (b) symptomatic leaves.
Figure 10.
Infection of Xcc CREA OF 1231.20 in B. oleracea var. botrytis; (a) non-inoculate control; (b) symptomatic leaves.


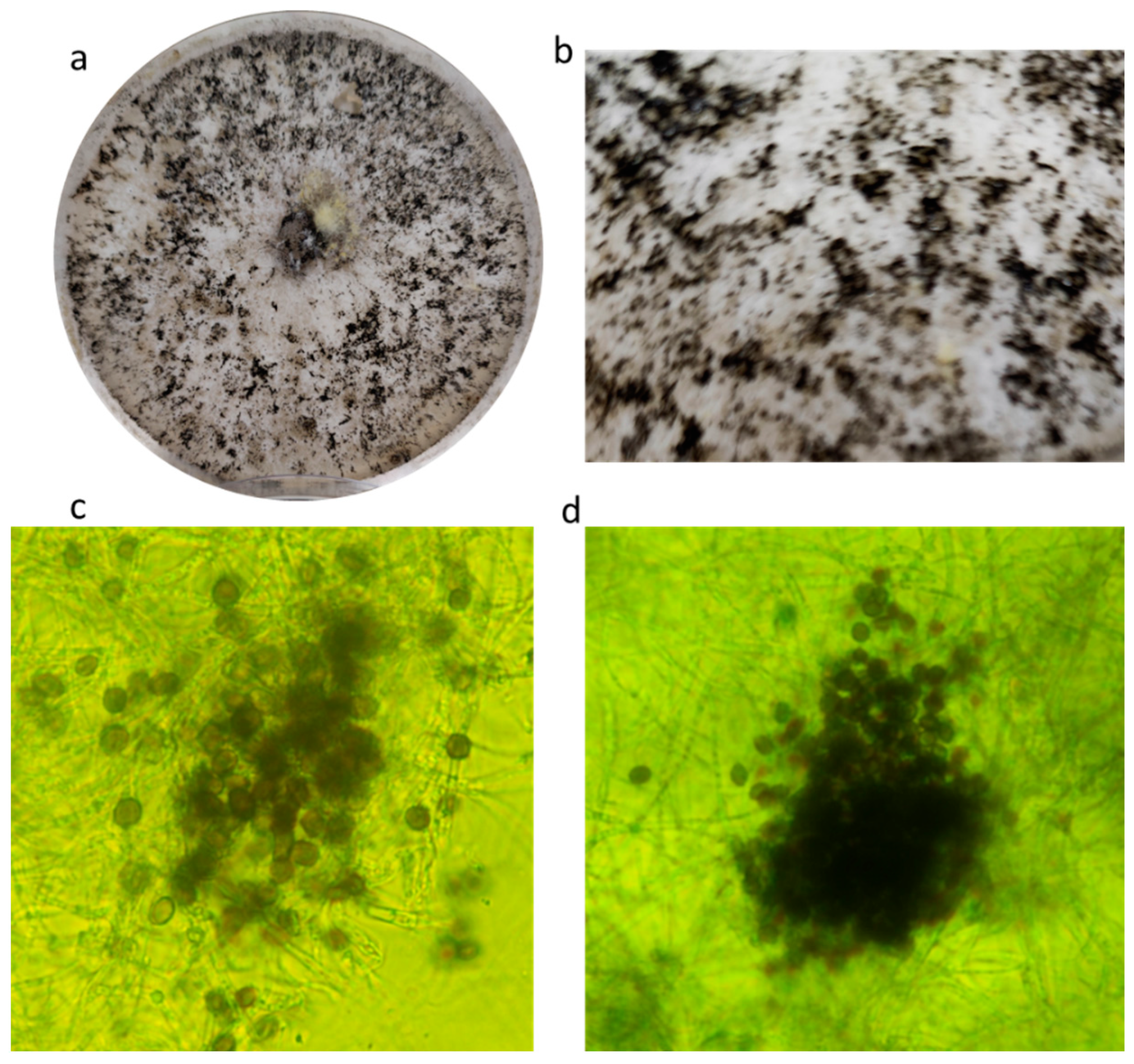{kind=link}
{kind=link}
{kind=link}
{kind=link}
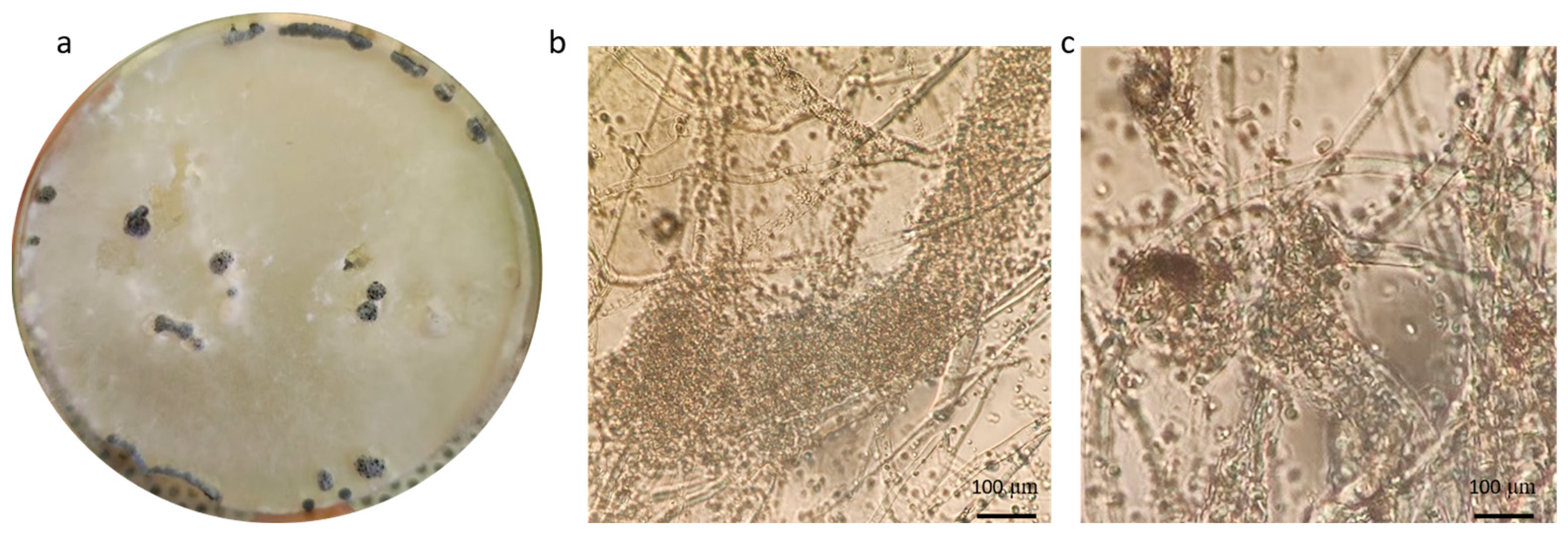{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of accessions subjected to surveillance for seed health. Species, accession name, geographic area of cultivation, fungi, bacteria, and viruses searched.
Table 1.
List of accessions subjected to surveillance for seed health. Species, accession name, geographic area of cultivation, fungi, bacteria, and viruses searched.
| Species | Accession | Geographic Area of Cultivation | Multiplication Field | Target Fungi (a) | Target Bacteria (b) | Target Viruses (c) | Analytical Methods (To Detect Pathogens (a, b, c)) | Literature |
|---|---|---|---|---|---|---|---|---|
| A. cepa | Agostina | Agro acerrano-mariglianese (NA) 1 | in situ | Botrytis spp. Alternaria porri Stemphylium vesicarium | / | / | (a) Mycological screening (agar plate method) | [20] |
| di Alife (alifana) | Alife (CE) 2 | ex situ | ||||||
| di Eremiti | Eremiti (SA) 3 | in situ | ||||||
| A. sativum | Bianco locale | Avellinese | in situ | Fusarium spp. Botrytis spp. Macrophomina phaseolina Sclerotinia spp. | Erwinia carotovora subsp. carotovora | / | (a) Mycological screening (agar plate method) (b) Isolation on Nutrient agar | [21,22] |
| ecotipo locale di Caposele | Caposele (AV) 4 | in situ | ||||||
| Brassica oleracea var. italica | Broccolo del Vallo di Diano | Vallo di Diano (NA) | ex situ | Alternaria brassicae, A. brassicicola, A. japonica, Phoma lingam Rizoctonia solani Sclerotinia spp. | Xanthomonas campestris pv. campestris and | / | (a) Mycological screening (agar plate method) (b) ISTA 7-019a (dilution plating method) | [20,23] |
| Broccolo dell’Olio | Agro nocerino-sarnese (SA) | ex situ | ||||||
| Broccolo di Natale | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Broccolo San Pasquale | Agro acerrano-mariglianese (NA) | ex situ | ||||||
| Brassica oleracea var. sabellica | Torzella riccia | Agro acerrano-mariglianese (NA) | ex situ | |||||
| Brassica rapa var. rapa | Catozza | Casertano, napoletano and Agro nocerino-sarnese (SA) | ex situ | Alternaria brassicae, A. brassicicola, A. japonica, Phoma lingam Rizoctonia solani Sclerotinia spp. | Xanthomonas campestris pv. campestris | / | (a) Mycological screening (agar plate method) (b) ISTA 7-019a (dilution plating method) | |
| L. sativus | Alta Irpinia | Avellinese | in situ | Ascochyta sp. Fusarium spp. Botrytis spp. | Pseudomonas spp. | / | (a) Mycological screening (agar plate method) (b) Dilution plating method | [24] |
| di Calitri | Calitri (AV) | ex situ | ||||||
| di Colliano | Colliano (SA) | ex situ | ||||||
| di Montefalcone | Montefalcone (BN) 5 | in situ | ||||||
| L. cicera | Maracuocciolo/Maracuccia | Ascea/Camerota (SA) | ex situ | Ascochyta sp. Fusarium spp. Botrytis spp. | Pseudomonas spp. | / | (a) Mycological screening (agar plate method) (b) Dilution plating method | [24] |
| L. culinaris | di Colliano | Calitri (AV) | in situ | Ascochyta sp. Fusarium spp. Botrytis spp. Sclerotinia spp. Rizoctonia solani | / | / | (a) Isolation on PDA | [24,25] |
| P. vulgaris | Bianco di Villa Santa Croce | Villa di Santa Croce (CE) | ex situ | Colletotrichum lindemuthianum Fusarium spp. Ascochyta phaseolorum Rizoctonia solani Macrophomina phaseolina Sclerotinia sclerotiorum Botrytis spp. | Pseudomonas savastanoi pv. phaseolicola Pseudomonas syringae pv. syringae Xanthomonas axonopodis pv. phaseoli | AMV CMV BCMV BCMNV | (a) Mycological 1 screening (agar plate method) (b) ISTA 7-21; ISTA 7-023 (Dilution plating method) (c) DAS ELISA (EPPO Standard PM 7/125(1)) | [23] |
| Butirro Acc. 2021 | Vico Equense (NA) | ex situ | ||||||
| Butirro Acc. 2020 | Vico Equense (NA) | ex situ | ||||||
| Cannellino Sessantino dei 30 anni | Acerra (NA) | in situ | ||||||
| Cannellino Bianco di Calitri | Calitri (AV) | in situ | ||||||
| dei 7 anni | Visciano (NA) | in situ | ||||||
| dei Signori | Agro nocerino-sarnese (SA) | ex situ | ||||||
| della Regina | Valle d’ll’Angelo (SA) | ex situ | ||||||
| della Regina acc.2 2021 Bianco | Montano Antilia (SA) | ex situ | ||||||
| della Regina Acc.2 2021 Gandolfi | Montano Antilia (SA) | ex situ | ||||||
| della Regina Acc.2 2021 Mazzamauro | Montano Antilia (SA) | ex situ | ||||||
| della Regina Acc.2 | San Lupo (BN) | ex situ | ||||||
| Della Regina di Gorga | Gorga (SA) | ex situ | ||||||
| della Regina | Montano Antilia (SA) | ex situ | ||||||
| Dente di Morto Acc. 2021 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.1 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.2 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.3 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.4 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.5 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.6 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc.7 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| Dente di Morto Acc. 2020 | Agro acerrano-mariglianese (NA) | in situ | ||||||
| di Prata Melizzano | Prata Sanni–a–Melizzano (BN) | ex situ | ||||||
| di Volturara Irpina | Volturara Irpina (NA) | in situ | ||||||
| Fasulo a tubbettiello | Ruviano (CE) | ex situ | ||||||
| Giallo del Fortore | Alto Fortore (BN) | ex situ | ||||||
| Lardari | Agerola (NA) | in situ | ||||||
| Regina | Grottaminarda (AV) | in situ | ||||||
| Rosso di Acerra | Acerra (NA) | ex situ | ||||||
| Schiacciatello | Cellole (SA) | ex situ | ||||||
| Tabaccanti | Vallo di Diano (NA) | ex situ | ||||||
| Tondino Bianco di Calitri | Calitri (AV) | in situ | ||||||
| Tondino di Castel di Sasso Acc. 2022 | Castel di Sasso (CE) | ex situ | ||||||
| Tondino di Villaricca | Villaricca (NA) | ex situ | ||||||
| Tondino di Villaricca | Villaricca (NA) | ex situ | ||||||
| Tondo Bianco | Sarno (SA) | ex situ | ||||||
| P. sativum | Centogiorni 2021 | Vesuvio/Agro nocerino-sarnese (NA) | ex situ | Ascochyta spp. Macrophomina phaseolina Rizoctonia solani Fusarium spp. | Pseudomonas syringae pv. pisi | CMV | (a) Mycological screening (agar plate method) (b) ISHI-Veg (Di-lution plating method) (c) DAS ELISA (EPPO Standard PM | [26] |
| Centogiorni Acc. 2021 | Acerra (NA) | ex situ | ||||||
| Centogiorni Acc. 2021 | Pollenatrocchia (NA) | in situ | ||||||
| Centogiorni Somma Vesuviana Acc. 2021 | Somma Vesuviana (NA) | in situ | ||||||
| Santacroce | Flegrea Area (NA) | in situ | ||||||
| Santacroce Acc. M 2021 | Flegrea Area (NA) | ex situ | ||||||
| Santacroce Acc. N 2021 | Flegrea Area (NA) | ex situ | ||||||
| Santacroce Acc. T 2021 | Napoletano | ex situ | ||||||
| V. faba | A sciabola Acc. 2021 | Agro acerrano-mariglianese (NA) | in situ | Ascochyta fabae Rizoctonia solani | Pseudomonas spp. | CMV | (a) Mycological screening (agar plate method) (b) Dilution plating method (c) DAS ELISA (EPPO Standard PM 7/125(1)) | [24] [27] |
| Di Colliano | COLLIANO (SA) | ex situ | ||||||
| lunga | Somma Vesuviana (NA) | ex situ | ||||||
| lunga Acc. 2020 | Nocerino-sarnese (NA) | ex situ | ||||||
| lunga Acc. 2021 | Nocerino-sarnese (NA) | ex situ | ||||||
| A Corna | Agro acerrano-mariglianese (NA) | ex situ | ||||||
| V. unguiculata | Corna dei Signori | Castel S. Giorgio (SA) | in situ | Colletotrichum sp., Fusarium spp. Rizoctonia solani Macrophomina phaseolina Sclerotinia sclerotiorum | Pseudomonas spp. | CMV, BCMV | (a) Mycological screening (agar plate method) (b) Dilution plating method (c) DAS ELISA (EPPO Standard PM 7/125(1)) | [27] |
| Corna dei signori (San Marzano) | San Marzano sul Sarno (SA) | ex situ | ||||||
| Fagiolino lungo San Marzano | San Marzano sul Sarno (SA) | in situ |
1: Naples; 2: Caserta, 3: Salerno; 4: Avellino; 5: Benevento.
Table 2.
GenBank records of CREA OF isolates.
| Accession Number | Locus | Strain Code | Species | Host/Species–Landrace |
|---|---|---|---|---|
| OQ280977 | ITS | CREA OF 1115.1 | Alternaria alternata | Allium cepa–“Agostina” |
| OQ132873 | ITS | CREA OF 1128.1 | Apiospora arundinis | P. vulgaris–“Tondino di Castel di Sasso” |
| OQ133464 | ITS | CREA OF 1113.1 | Sclerotinia sclerotiorum | V.–faba–“Lunga (nocerino-sarnese) |
| OQ134131 | ITS | CREA OF 190.4 | Fusarium oxysporum | L. culinaris–“di Colliano” |
| OQ134394 | ITS | CREA OF 1231.3 | Alternaria brassicicola | B. oleracea var. sabellica–“Torzella riccia” |
| OQ148383 | ITS | CREA OF 1231.1 | Alternaria alternata | B. oleracea var. sabellica–Torzella riccia” |
| OQ148397 | ITS | CREA OF 1231.5 | Alternaria alternata | B. oleracea var. sabellica– “Torzella riccia” |
| OQ148398 | ITS | CREA OF 1231.4 | Alternaria alternata | B. oleracea var. sabellica– “Torzella riccia” |
| OQ148399 | ITS | CREA OF 1231.6 | Alternaria alternata | B. oleracea var. sabellica– “Torzella riccia” |
| OQ148403 | ITS | CREA OF 1235.2 | Ascochyta fabae | V. faba–“A Corna” |
| OQ148428 | ITS | CREA OF 1122.4 | Ascochyta pisi | P. sativum –“Centogiorni (Pollenatrocchia)” |
| OQ148433 | ITS | CREA OF 1122.5 | Ascochyta pisi | P. sativum–“Santacroce Acc.M” |
| OQ148577 | ITS | CREA OF 1119.1 | Fusarium proliferatum | A. sativum–“Bianco locale” |
| OQ148588 | ITS | CREA OF 355.4 | Stemphylium globuliferum | L. cicera–“Maracuoccio” |
| OQ249627 | β-tub | CREA OF 1128.1 | Apiospora arundinis | P. vulgaris–“Tondino di Castel di Sasso” |
| OQ249628 | TEF 1-α | CREA OF 1113.1 | Sclerotinia sclerotiorum | V. faba–“Lunga (nocerino-sarnese) |
| OQ249629 | TEF 1-α | CREA OF 190.4 | Fusarium oxysporum | L. culinaris–“Di Colliano” |
| OQ249630 | TEF 1-α | CREA OF 1112.1 | Stemphylium vesicarium | V. faba–“A Sciabola” |
| OQ266773 | ITS | CREA OF 1112.1 | Stemphylium vesicarium | Vicia faba -A sciabola |
| OQ302538 | ITS | CREA OF 1231.2 | Alternaria infectoria | B. oleracea var. sabellica– “Torzella riccia” |
Table 3.
Primer pairs used to characterize the isolates.
| Locus/Target | Definition | Primers | Sequence (5′-3′) | Reference | Isolates |
|---|---|---|---|---|---|
| EF-1α | Translation elongation factor 1α | EF1-728F | CATCGAGAAGTTCGAGAAGG | [28] | CREA OF 1113.1 CREA OF 1112.1 |
| EF1-968R | TACTTGAAGGAACCCTTACC | ||||
| EF1-688F | CGGTCACTTGATCTACAAGTGC | [29] | CREA OF 190.4 | ||
| EF1-1251R | CCTCGAACTCACCAGGTACCG | ||||
| ITS1 | Internal transcribing spacer 1 | ITS1 | TCCGTAGGTGAACCTGCGG | [30] | All |
| ITS4 | TCCTCCGTCTATTGATATGC | ||||
| β-tub | β-tubulin | T1 | AACATGCGTGAGATTGTAAGT | [31] | CREA OF 1128.1 |
| β-tub-2B | ACCCTCAGTGTAGTGACCCTTGGC | ||||
| Calmodulin/Macrophomina spp. | Calmodulin | MpCalF | CAATCTCTTTCTCCCCTACAGGA | [32] | CREA OF 1128.3 |
| MpCalR | ACTGCGCAAAAGCGCCAGTAAAC | ||||
| MsCalF | CAATGTCTTTCTCCACTGCAGGA | ||||
| MsCalR | TACTGCGCCAAAGCGGCAGTAAA | ||||
| MeCalF | AGCCCGCCTTGCTCCACCCTGTTCT | ||||
| MeCalR | TGTATACTGCGCAAAAGCGGCAGT | ||||
| EF-1α | Translation elongation factor 1α | MpTefF | AAACACACTTTTCGCACTCCTGC- | ||
| MpTefR | TATGCTCGCAGAGAAGAACACGA | ||||
| MsTefF | GGCACACTTTTCGCGCTTCTGTA | ||||
| MsTefR | TGTGCTCGCTGGGAAGAACATGA | ||||
| MeTefF | AAGCATACTTTTCGTGCTCCTGC | ||||
| MeTefR | AAAGGAACATGAGTGGCCAAAAA | ||||
| Xap/Xff | p7X4c | GGCAACACCCGATCCCTAAACAGG | [23] | CREA OF 522, 539, 540, 541, 542, 543, 544, 545, 546 | |
| p7X4e | CGCCGGAAGCACGATCCTCGAAG | ||||
| Xcc | DLH120 | CCGTAGCACTTAGTGCAATG | [23] | CREA OF 1231.20, 1231.21, 1231.22 | |
| DLH125 | GCATTTCCATCGGTCACGATTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dell’Olmo, E.; Zaccardelli, M.; Onofaro Sanaja, V.; Basile, B.; Sigillo, L. Surveillance of Landraces’ Seed Health in South Italy and New Evidence on Crop Diseases. Plants 2023, 12, 812. https://doi.org/10.3390/plants12040812
AMA Style
Dell’Olmo E, Zaccardelli M, Onofaro Sanaja V, Basile B, Sigillo L. Surveillance of Landraces’ Seed Health in South Italy and New Evidence on Crop Diseases. Plants. 2023; 12(4):812. https://doi.org/10.3390/plants12040812
Chicago/Turabian StyleDell’Olmo, Eliana, Massimo Zaccardelli, Vincenzo Onofaro Sanaja, Boris Basile, and Loredana Sigillo. 2023. "Surveillance of Landraces’ Seed Health in South Italy and New Evidence on Crop Diseases" Plants 12, no. 4: 812. https://doi.org/10.3390/plants12040812
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.