
Alternaria arborescens and A. italica Causing Leaf Blotch on Celtis julianae in China
by
 , and
, and
Yang-Chun-Zi Liao
1,2,†,
Yi-Jia Cao
1,2,†,
Yu Wan
1,2,
Hui Li
1,2,
De-Wei Li
3,* and
Li-Hua Zhu
1,2,* 1
College of Forestry, Nanjing Forestry University, Nanjing 210037, China
2
Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
3
The Connecticut Agricultural Experiment Station Valley Laboratory, Windsor, CT 06095, USA
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2023, 12(17), 3113; https://doi.org/10.3390/plants12173113
Submission received: 10 August 2023
/
Revised: 25 August 2023
/
Accepted: 28 August 2023
/
Published: 30 August 2023
(This article belongs to the Section Plant Protection and Biotic Interactions)
Abstract
:Celtis julianae Schneid. is widely planted as a versatile tree species with ecological and economic significance. In September 2022, a leaf blotch disease of C. julianae was observed in Nanjing, Jiangsu, China, with an infection incidence of 63%. The disease led to severe early defoliation, significantly affecting the ornamental and ecological value of the host tree. The accurate identification of pathogens is imperative to conducting further research and advancing disease control. Koch’s postulates confirmed that the fungal isolates (B1–B9) were pathogenic to C. julianae. The morphology of the characteristics of the pathogen matched those of Alternaria spp. The internal transcribed spacer region (ITS), large subunit (LSU) and small subunit (SSU) regions of rRNA, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), Alternaria major allergen gene (Alt a 1), RNA polymerase second largest subunit (RPB2), and portions of translation elongation factor 1-alpha (TEF1-α) genes were sequenced. Based on multi-locus phylogenetic analyses and morphology, the pathogenic fungi were identified as Alternaria arborescens and A. italica. The findings provided useful information for disease management and enhanced the understanding of Alternaria species diversity in China. This is the first report of A. arborescens and A. italica causing leaf blotch of C. julianae in China and worldwide.
1. Introduction
Celtis julianae Schneid. (Ulmaceae) is a large deciduous tree that can reach a height of over 25 m. It serves as a valuable landscaping and shelter tree species due to its ability to withstand soot and toxic gases [1]. Additionally, its kernels are used as raw material for soap and lubricating oil production due to their high oil content. The numerous hairs on leaf surfaces enable it to trap airborne particulates, contributing to air purification [2]. In September 2022, a leaf blotch disease was observed on C. julianae at Nanjing Forestry University (118°48′26″ E, 32°4′52″ N). The disease led to early defoliation in severe cases, resulting in a reduction of its ornamental and ecological value.
The Alternaria genus comprises approximately ca. 382 species separated into 29 sections [3,4,5,6]. Alternaria species are crucial invasive pathogen that can colonize a wide range of hosts, including various plants in the phyllosphere, and animals, including humans [3,7,8,9,10]. Over 4000 monocotyledonous and dicotyledonous plant species are affected by Alternaria spp. [3,6,11,12,13]. For example, A. alternata (Fr.) Keissler is known to cause leaf spots on Prunus salicina [14]. In Oman, several species of Alternaria are associated with leaf spots on date palm and wheat produce, leading to reduced market value and significant economic losses [15]. Additionally, A. arborescens E.G. Simmons has been reported to cause leaf spots on Pereskia aculeata in Brazil [16]. In addition, the conidia of Alternaria are the most common airborne allergens and have been determined to be significant triggers of allergic rhinitis and allergic asthma [17,18]. Furthermore, Alternaria species have emerged as important human invasive pathogens in immune-compromised patients [19,20]. Thus, the Alternaria species deserves further studies.
The traditional identification of plant pathogenic fungi has mainly relied solely on morphological characteristics and host association. However, when relying solely on morphological characteristics, such as conidia, conidiophores, conidiogenous cells, and fruiting body, the pathogenic fungi may not be accurately classified and determined. Phylogenetic analyses have been widely applied for decades, leading to the discovery of numerous new species [21,22,23]. Initially, the internal transcribed spacer region (ITS) was commonly used for taxonomic purposes. However, studies have pointed out the limitations of using ITS sequence data [24,25,26]. In many fungal genera, the ITS locus only resolves taxa to the genus level due to the intraspecific and even intragenomic polymorphisms commonly observed [27,28]. Moreover, a substantial percentage of ITS sequences in GenBank are derived from misidentified specimens or cultures [29,30]. As a result, secondary DNA barcodes have been proposed for various genera of plant pathogenic fungi [31,32,33]. Nowadays, multi-locus phylogenetic analysis is considered more appliable and reliable in fungal taxonomy [34].
Similarly, Alternaria species were traditionally classified based on morphological characteristics of their reproductive structures and sporulation patterns under various conditions [35]. Alternaria spp. are dematiaceous fungi, exhibiting grey-olive/brown colonies. The conidia of Alternaria are dictyospores. These can either be found alone or in chains [36]. However, the subtle differences between Alternaria spp. and the morphological variation under different conditions can make identification based on solely on morphological characteristics challenging. With the advent of molecular analyses, researchers have examined phylogenetic relationships among Alternaria species. Lawrence et al. [12,18] first determined 27 sections in Alternaria through multi-locus phylogeny. Woudenberg et al. [6] used six loci to establish 24 sections, including 16 newly described sections. By combining seven loci, a consensus phylogeny was generated. Ghafri et al. [37] introduced Alternaria section Omanenses in 2019, and Gannibal et al. [38] introduced Alternaria section Helianthiinficiens in 2022, bring the total number of Alternaria species to 29 sections. Currently, a clear and stable phylogenetic classification has been established at the species level among Alternaria spp., providing a reliable basis for this study.
The objectives of this study were (1) to isolate and identify the pathogen of leaf blotch on C. julianae according to morphological characteristics and multilocus phylogenetic analyses, and (2) to confirm the pathogenicity of the isolates on C. julianae.
2. Results
2.1. Disease Symptoms and Fungal Isolations
The leaf blotch disease was observed on C. julianae at the campus of Nanjing Forestry University, China, with an incidence of 63% (63/100 plants). On average, 80% of the leaves per tree showed the presence of this leaf blotch disease (Figure 1A,B). The symptoms initially appeared as small, light brown spots. Measuring 1–2 mm in diameter, they were surrounded by yellow halos (Figure 1C). Subsequently, the spots gradually expanded into irregular necrotic blotches with dark brown borders, and the halos developed around the dark brown borders, leading to an increase in the size of the blotches over time (Figure 1D). Eventually, the blotches coalesced into large necrotic areas, resulting in leaf shrinkage, wrinkling, and collapse (Figure 1E). Numerous brown conidial chains were observed on the lesions of infected leaves (Figure 1F,G).
Diseased leaves were collected twice for isolating fungal pathogens in September and in October 2022, respectively. In the September experiment, three types of colonies were found with a frequency of 63%, 24%, and 13%, respectively. Similarly, in the October experiment, three types of colonies were found with a frequency of 54%, 38%, and 8%, respectively. According to the ITS sequence alignment, the three types of colonies belonged to the genera Nigrospora, Alternaria and Nothophoma, respectively (Table 1).
2.2. Pathogenicity Tests
Five days post-inoculation in vitro, all detached leaves inoculated with nine isolates (B1–B9) of Alternaria spp. appeared to display brown spot symptoms at the inoculation sites, whereas leaves inoculated with Nigrospora sp., Nothophoma sp. and control leaves did not show any symptoms. Subsequently, in vivo experiments were conducted that used three representative fungal isolates (B1, B2 and B3) of the Alternaria species to inoculate healthy C. julianae seedlings. Seven days after inoculation, the inoculated leaves appeared to be brown spots with yellow halos, resembling the early symptoms observed on the leaves infected in the field. In contrast, the control leaves remained healthy (Figure 2). The same fungus was re-isolated from the lesions, and no other fungi were isolated from the control leaves. This fulfills Koch’s postulates, confirming that isolates B1, B2 and B3 are the causal agents of leaf blotch on C. julianae.
2.3. Multigene Phylogenetic Analyses
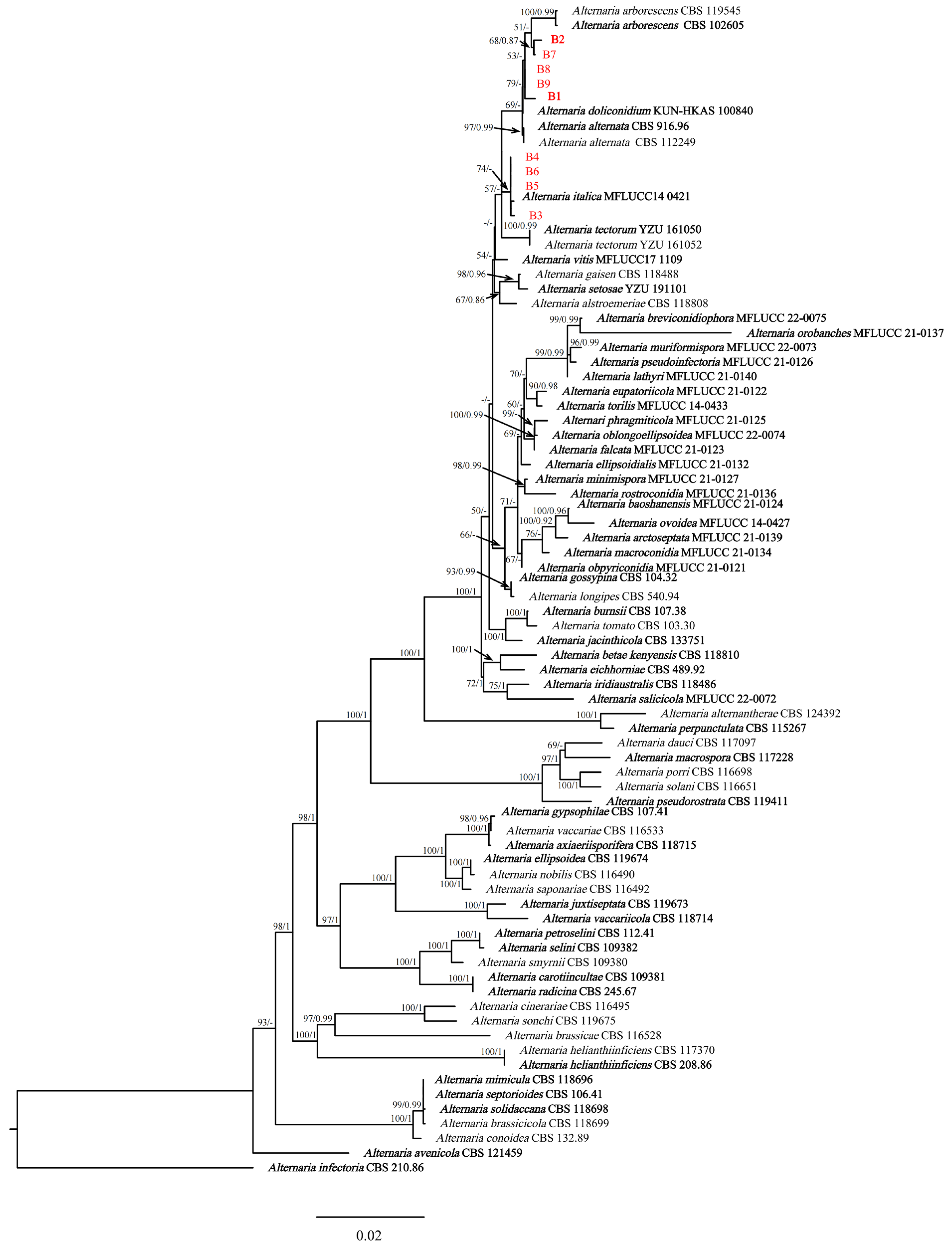
Phylogenetic analyses performed using maximum-likelihood and Bayesian inference techniques placed the four isolates (B3, B4, B5, and B6) in the same cluster with Alternaria italica J.F. Li, Camporesi & K.D. Hyde (extype: MFLUCC 14-0421T). Meanwhile, there were five isolates (B1, B2, B7, B8, and B9) in the same cluster with A. arborescens (extype: CBS 102605) (Figure 3). The phylogenetic trees generated using ML and BI methods showed consistent topology. Based on the phylogenetic analyses conducted using the concatenated sequences of seven genes/regions (ITS, LSU, SSU, GAPDH, Alt a 1, RPB2 and TEF1-α), B1, B2, B7, B8, and B9 were identified as A. arborescens, while B3, B4, B5, and B6 were determined to be A. italica.
2.4. Morphology and Taxonomy
- Alternaria arborescens E.G. Simmons (Figure 4)
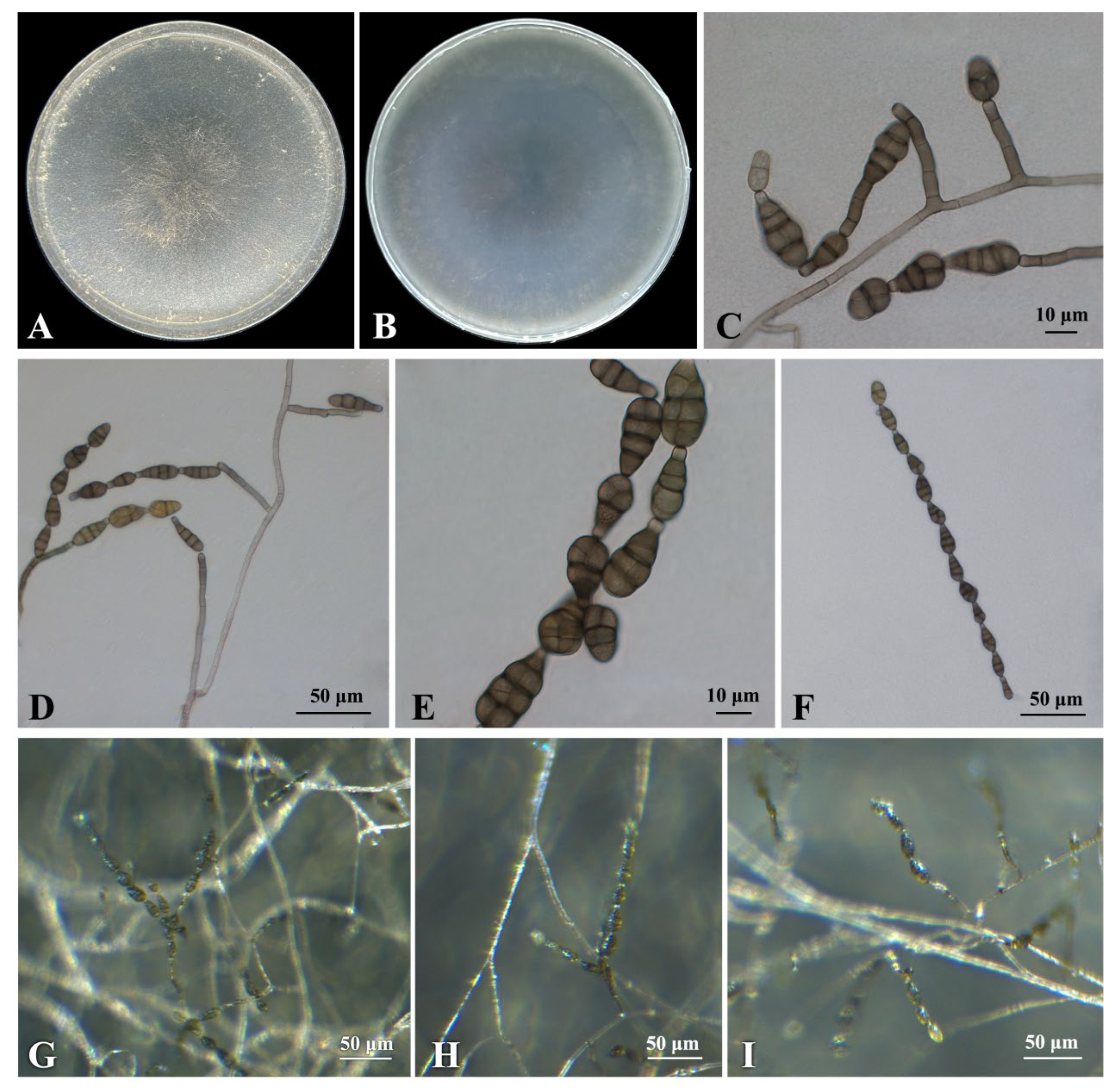
Culture characteristics: On potato–carrot agar (PCA) and V8 agar (V-8), colonies of isolate B1 were circular, flat, and granulated with undulating edges. The colony was grayish green, and the reverse side was greenish brown (Figure 4A,B).
Description: Sexual morph not observed. Under the Zeiss Axio Imager A2m microscope (Carl Zeiss AG, Oberkochen Germany), the hyphae were hyaline to light brown, septate, and (3.0−)3.7 − 5.3(−7.1) μm (mean ± SD = 4.5 ± 0.8 μm, n = 30) wide. Conidiophores were solitary, dark brown, straight or curved, 2–8 septa, and variable in length, (32.7−)41.5 − 94.3(−121.6) × (3.2−)3.7 − 4.7(−5.3) μm (mean ± SD = 67.9 ± 26.4 × 4.2 ± 0.5 μm, n = 30) (Figure 4C,D). Conidiogenous cells (5.5−)6.2 − 9.2(−11.8) × (3.4−)4.0 − 4.8(−5.0) μm (mean ± SD = 7.7 ± 1.5 × 4.4 ± 0.4 μm, n = 30). Conidia were oval or obclavate, brown to dark brown, with 1–4 transverse septa and 1–4 longitudinal or oblique septa, constricted at the septa, (19.7−)23.6 − 28.4(−34.3) × (9.8−)11.0 − 12.8(−13.7) μm (mean ± SD = 26.0 ± 2.4 × 11.9 ± 0.9 μm, n = 30) (Figure 4E). The beaks (2.7−)3.3 − 5.1(−6.4) × (2.8−)3.4 − 4.4(−4.9) μm (mean ± SD = 4.2 ± 0.9 × 3.9 ± 0.5 μm, n = 30). Conidial chains were simple or branched with 1–15 conidia. (Figure 4F).
Specimens examined: China, Jiangsu province, Nanjing city, isolated from leaves of Celtis julianae, 1 September 2022, Yijia Cao, cultures: CFCC 59038 (=B1), CFCC 59039 (=B2), B7, B8 and B9.
Notes: The phylogenetic analyses showed that five isolates (B1, B2, B7, B8 and B9) were in a clade with A. arborescens (Figure 3). The morphological features of the five isolates matched those of A. arborescens [39]. Based on the morphology and phylogeny, B1, B2, B7, B8 and B9 were identified as A. arborescens.
- Alternaria italica J.F. Li, Camporesi & K.D. Hyde (Figure 5)
Culture characteristics: On PCA and V-8 media, the colony appears flat with gray margin. It is greyish green and cottony, covering the Petri dish after 7 days and showing abundant sporulation. The reverse side is dark greyish-green and radial (Figure 5A,B). After a week, the culture has increased the amount of white aerial mycelium in the center of the plate.
Description: Sexual morph not observed. The hyphae were colorless, hyaline to light brown, septate, (3.2−)3.6 − 5.0(−5.7) μm (mean ± SD = 4.3 ± 0.7 μm, n = 30) wide. Conidiophores macronematous, mononematous, flexuous or sigmoid, 0–5 septate, simple or branched, smooth, and hyaline to light brown, (13.5−)20.1 − 36.7(−46.2) × (3.1−)3.8 − 4.8(−5.3) μm (mean ± SD = 28.4 ± 8.3 × 4.3 ± 0.5 μm, n = 30) (Figure 5D,E). Lateral secondary conidiophores were observed but relatively uncommon. Conidiogenous cells were at the tip of conidiophores, (4.8−)5.8 − 8.0(−10.1) × (3.4−)3.8 − 4.8(−6.2) μm (mean ± SD = 6.9 ± 1.1 × 4.3 ± 0.5 μm, n = 30). Conidial chains were commonly single file, occasionally branched with 2–8 conidia (Figure 5C,D). Conidia dictyospores, pale brown to brown, variable in size and shape, but often obclavate to obpyriform, with up to 8 transverse and usually 0–2 longitudinal or oblique septa, slightly constricted at the septa, (27.7−)29.6 − 40.0(−51.3) × (7.6−)8.2 − 10.8(−13.1) μm (mean ± SD = 34.8 ± 5.2 × 9.5 ± 1.3 μm, n = 30) (Figure 5C). The apex of the conidia bears a beak, pale brown, most of the beaks are relatively short and well rounded, (3.1−)4.0 − 8.2(−12.2) × (3.1−)3.5 − 4.3(−5.1) μm (mean ± SD = 6.1 ± 2.1 × 3.9 ± 0.4 μm, n = 30).
Specimens examined: China, Jiangsu province, Nanjing city, isolated from leaves of Celtis julianae, 1 September 2022, Yijia Cao, cultures: CFCC 59359 (=B3), CFCC 59309 (=B4), CFCC 59310 (=B5), and CFCC 59311 (=B6).
3. Discussion
The genus Alternaria was first described by Nees von Esenbeck (1816), with A. tenuis as the type of species. Historically, the identification and classification of Alternaria species heavily relied on measurements and descriptions of morphological characteristics [41], complemented later by molecular phylogeny [42,43]. However, recent studies have challenged the morphological basis for the identification of some species in Alternaria [11,13,44]. Currently, the genus Alternaria contains 29 sections, and the main morphological characteristic of Alternaria sect. Alternaria is the production of short conidia in chains [6,18,45]. Woudenberg et al. [6] established species in Alternaria sect. Alternaria based on ITS. Subsequent whole-genome sequencing, transcriptome comparisons, and multi-gene sequencing further rearranged Alternaria sect. Alternaria into 11 phylogenetic species and one species complex [13]. Following a number of studies conducted by Li et al. [3,4], Cannibal et al. [45,46], Wanasinghe et al. [47], Jayawardena et al. [48] and Nishikawa et al. [49], currently over 89 species constitute the sect. Alternaria. Traditional morphological methods with molecular phylogeny are vital for identifying Alternaria species. In this study, a multi-locus phylogeny based on a concatenated ITS, LSU, SSU, GAPDH, Alt a 1, RPB2 and TEF1-α sequences, combined with morphological characteristics, revealed two known species, A. arborescens and A. italica, within the sect Alternaria. Additionally, our study also reported new host records for A. arborescens and A. italica. It is worth noting that many pathogenic fungal species are named after their hosts, and that the two Alternaria species identified in this study belong to the same section and is isolated from the same host. The similarities in their morphological features highlight the importance of taking cautious approaches in identifying and naming Alternaria species.
Alternaria is among the primary mycotoxigenic fungal genera found in cereals worldwide [50]. It can produce a variety of mycotoxins derived from secondary metabolism, with about 70 toxic metabolites described so far [35]. Some pathogenic Alternaria spp. utilize a diverse array of pathogenic toxins to infect plant tissues [51,52]. Certain species of Alternaria produce host-specific toxins (HSTs) that influence their pathogenicity and virulence. For example, the Japanese pear pathotype associated with Alternaria produces AK toxins I and II, which only exhibit toxicity in susceptible pear cultivars [35]. In addition, HSTs affect the taxonomy of the genus as fungal systematics include the analysis of secondary metabolites, providing information for use in species differentiation. At present, several researchers have employed chemotaxonomy and pathogenicity characteristics to assist with classification [53,54,55]. However, the method cannot be used independently for classification, because members within the same group may not share a common metabolite profile [50]. Therefore, the application of polyphasic approaches could provide strong support in fungal taxonomy, and further studies are warranted.
In our study, we observed that the isolation rates of Nigrospora spp. were consistently higher than those of Alternaria spp. in two independent fungal isolations. However, coexistence observation and pathogenicity testing showed that Alternaria spp. was the pathogen causing leaf blotching of C. julianae. Previous studies have indicated that species the of genus Nigrospora possess a strong saprobic nature and commonly occur as plant endophytes or saprobes on different hosts [56,57]. Nigrospora spp. may have more advantages in competing for nutrients in substrates compared to Alternaria spp., which can slow the growth of Alternaria. This could be the reason for the higher isolating rates of Nigrospora than those of Alternaria spp.
Celtis julianae is an important ornamental species mainly found in the temperate biome, with its native range overlaying the central and southern parts of China [58]. It has a high level of wind resistance due to its deep roots, making it an ideal species for embankment protection and water conservation [59]. Additionally, C. julianae helps maintain air humidity, reduce heat, and create various shadows with its large and dense leaves [60]. Economically, C. julianae has rich bark fiber, which can be used for making paper, and the core of its fruit can be used in oil and soap manufacture. In summary, C. julianae is a multi-functional species with ornamental, ecological and economic value. Currently, research on diseases affecting C. julianae is insufficient. Therefore, the investigation of leaf blotch on C. julianae and identification of pathogens have significance in providing the theoretical basis for reducing the economic and ecological losses caused by this disease.
4. Materials and Methods
4.1. Sampling and Isolation of Fungi
From September to October 2022, the isolation of fungi was conducted twice. Each time, 25 symptomatic leaves were collected from three infected plants on the campus of Nanjing Forestry University. The symptomatic leaves were first rinsed under running water and dried on sterilized filter paper. One-hundred small tissue (3 × 3 mm) samples were cut from lesion margins and surface-sterilized in 75% ethanol for 30 s, followed by the use of 1% NaClO for 90 s. The samples were then rinsed 3 times in sterile water, dried on sterilized filter paper, and finally plated onto potato dextrose agar (PDA) supplemented with ampicillin. The plates were incubated at 25 °C in an incubator, MIR-553 (Sanyo, Osaka Japan) [61]. After 3 days, the growing edges of the mycelium from the plant materials were cut and transferred to fresh PDA media to obtain a pure culture [62].
4.2. Pathogenicity Tests
To determine the pathogenicity of the isolates, experiments were conducted on both detached leaves and attached leaves. Healthy leaves from the field were collected, rinsed under tap-water for 15 min, and then wounded with sterile needles after drying on sterilized filter paper. For each type of test, two isolates were selected. For inoculation, plugs (5 mm diam.) were cut out from the growing edges of 5-day-old cultures and placed onto the wounds. The PDA plugs were used as controls. Five wounded leaves were treated with isolate and control plug. After inoculation, the leaves were placed into Petri dishes to maintain their humidity and kept at 25 °C in an incubator. Subsequently, nine representative fungal isolates (B1–B9) were selected from pathogenic species for in vivo inoculation with mycelial plugs. To confirm Koch’s postulates, further pathogenicity tests were performed by inoculating conidial suspensions on leaves of C. julianae seedlings that were wounded with a sterile needle. The leaves were inoculated with the three representative isolates (B1–B3) using 10 µL of conidial suspensions (106 conidia/mL). Five plants were used for each treatment, and three leaves of each plant were inoculated. The control was treated with 10 µL of sterile water. All inoculated seedlings were covered with plastic bags, and sterilized water was sprayed into the bags daily to maintain a moist microclimate. All seedlings were kept in a growing chamber at 20 ± 2 °C/10 ± 2 °C (day/night) and observed regularly. Leaves that showed typical symptoms after the inoculation were used for re-isolations.
4.3. DNA Extraction and PCR Amplification
Nine isolates were cultured on potato dextrose agar (PDA), V8 agar (V-8) and potato–carrot agar (PCA) media at 25 °C in a constant temperature incubator with a 12/12 h light/dark cycle. Morphological identification was based on colony morphology and characteristics of conidia, conidiophores, conidiogenous cells, such as the shape and color of the colony, as well as the shape, size, color, septation, and presence of a beak in the conidia. A Zeiss Axio Imager A2m microscope (Carl Zeiss AG, Oberkochen Germany) and Zeiss stereo microscope (SteRo Discovery v20) were used to observe, describe, and measure conidia and other structures (n = 30).
4.4. Morphological Identification
For accurate pathogen identification, the genomic DNA was extracted using a modified CTAB method [63]. The internal transcribed spacer (ITS) [64], large subunit (LSU) and small subunit (SSU) regions of rRNA [64,65], glyceraldehyde-3-phosphate dehydrogenase (GAPDH) [66], Alternaria major allergen gene (Alt a 1) [67], RNA polymerase second largest subunit (RPB2) [68] and portions of translation elongation factor 1-alpha (TEF1-α) [69] genes/region were sequenced using primers ITS1/4, LR0R/LR05, NS1/NS4, GPD1/GPD2, Alt-for/Alt-rev, RPB2-5F2/RPB2-7cR and EF1-728F/EF1-986R, respectively. PCR was carried out in a 50 µL reaction mixture containing 2 µL DNA; 2 µL of 10 µM primer, both forward and reverse (Table 2); 25 µL of Taq DNA polymerase mix; and 19 µL double-distilled water. The PCR amplifications were carried out with the following cycling parameters: initial denaturation at 94 °C for 3 min, followed by 33 cycles of 30 s at 94 °C, annealing at a suitable temperature for 30 s for different loci: 55 °C for ITS, 52.4 °C for LSU, 53 °C for SSU and RPB2, 59.5 °C for GAPDH, 60.5 °C for Alt a 1, extension at 72 °C for 30 s, and a final elongation at 72 °C for 10 min. The cycling parameter for TEF1-α was as follows: 94 °C for 5 min; 40 cycles at 94 °C for 30 s; 59 °C for 30 s; 72 °C for 45 s; and a final extension at 72 °C for 7 min. The PCR products were sequenced at Sangon Biotech Co., Ltd. (Nanjing, China).
4.5. Multigene Phylogenetic Analyses
The obtained sequences were analyzed using BLAST (https://www.ncbi.nlm.nih.gov/, accessed on 2 August 2023) to retrieve sequences with high similarities to the query sequences. Seventy-one isolates of 45 Alternaria species were obtained from GenBank for phylogenetic use in analyses (Table 3). The sequences of each gene/region were aligned with MAFFT ver. 7.313 (https://mafft.cbrc.jp/alignment/server, accessed on 2 August 2023) and manually adjusted using BioEdit ver. 7.0 [70,71]. The seven genes/region were concatenated by PhyloSuite ver. 7.313 [72]. ModelFinder was used to select the best-fit model, and phylogenetic relationships were inferred using maximum-likelihood (ML) analysis in IQtree ver. 1.6.8 and Bayesian inference (BI) in MrBayes 3.2.6 [73,74,75]. The ML analysis used the best model: GTR + I + G4 + F, with 1000 bootstrap replicates. The BI analysis used the best model: SYM + I + G, with 2 parallel runs and 2,000,000 generations, discarding the initial 25% of sampled data as burn-in. Phylogenetic trees were visualized using FigTree ver. 1.4.3 (https://tree.bio.ed.ac.uk/software/figtree/, accessed on 2 August 2023).
5. Conclusions
In the present study, we conducted fungal pathogen isolation and pathogenicity tests. We identified the nine fungal isolates (B1–B9) obtained, which are causing leaf blotch of C. julianae, based on multi-locus phylogenetic analyses using loci of ITS, LSU, SSU, GAPDH, Alt a 1, RPB2, TEF1-α, and morphological characteristics. The isolates were determined to be A. arborescens and A. italica within the Alternaria section Alternaria. To the best of our knowledge, this is the first report of A. arborescens and A. italica causing leaf blotch of C. julianae in either China or the wider world. The results of the study provide imperative and fundamental information for understanding the disease and performing future studies on the fungi/pathogens and the disease from mycological and phytopathological aspects. It is the first step in advancing the management of the disease.
Author Contributions
Y.-C.-Z.L., Y.-J.C., Y.W., H.L., L.-H.Z. and D.-W.L. designed the study and were involved in writing the paper; Y.-J.C. and L.-H.Z. were responsible for sample collections; Y.-J.C. was responsible for pathogenicity tests; Y.-C.-Z.L., Y.-J.C., H.L. and Y.W. were involved in morphological identification; Y.-C.-Z.L., Y.-J.C. and Y.W. were involved in phylogenetic analyses; L.-H.Z., D.-W.L., Y.-C.-Z.L., Y.-J.C. and Y.W. contributed to planning and editing of the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key R & D Program of China (2022YFD1401005), the National Natural Science Foundation of China, grant number 31971659.
Data Availability Statement
All data generated or analyzed during this study are included in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, X.D.; Liu, H.D.; Jing, S.; Liu, Y.H. The Complete Chloroplast Genome of Celtis julianae (Ulmaceae). Mitochondrial DNA Part B Resour. 2021, 6, 517–518. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, L.J. New Golden Leaf Cultivar of Celtis julianae (‘Golden Phoenix’) and Its Primary Research. HortSci. Publ. Am. Soc. Hortic. Sci. 2004, 39, 895. [Google Scholar] [CrossRef]
- Li, J.F.; Jiang, H.; Jeewon, R.; Hongsanan, S.; Bhat, D.J.; Tang, S.M.; Lumyong, S.; Mortimer, P.E.; Xu, J.C.; Camporesi, E.; et al. Alternaria: Update on Species Limits, Evolution, Multi-Locus Phylogeny, and Classification. Stud. Fungi 2023, 8, 1. [Google Scholar] [CrossRef]
- Li, J.F.; Phookamsak, R.; Jiang, H.; Bhat, D.J.; Camporesi, E.; Lumyong, S.; Kumla, J.; Hongsanan, S.; Mortimer, P.E.; Xu, J.C.; et al. Additions to the Inventory of the Genus Alternaria Section Alternaria (Pleosporaceae, Pleosporales) in Italy. J. Fungi 2022, 8, 898. [Google Scholar] [CrossRef]
- Gou, Y.N.; Aung, S.L.L.; Htun, A.A.; Huang, C.X.; Deng, J.X. Alternaria Species in Section Alternaria Associated with Iris Plants in China. Front. Microbiol. 2022, 13, 1036950. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria Redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Lee, H.B.; Jang, Y.S.; Cho, Y.R.; Kim, G.H.; Kim, J.J. Phylogenetic Analysis of Major Molds Inhabiting Woods. Part 4. Genus Alternaria. Holzforschung 2014, 68, 247–251. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J. Alternaria spp.: From General Saprophyte to Specific Parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Patriarca, A.; Magan, N. Alternaria in Food: Ecophysiology, Mycotoxin Production and Toxicology. Mycobiology 2015, 43, 93–106. [Google Scholar] [CrossRef]
- Wijayawardene, N.H.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and Fungus-like Taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Armitage, A.D.; Barbara, D.J.; Harrison, R.J.; Lane, C.R.; Sreenivasaprasad, S.; Woodhall, J.W.; Clarkson, J.P. Discrete Lineages within Alternaria alternata Species Group: Identification Using New Highly Variable Loci and Support from Morphological Characters. Fungal Biol. 2015, 119, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and Taxonomy of the Pleomorphic Genus Alternaria. Mycol. Prog. 2015, 15, 3. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria Section Alternaria: Species, Formae Speciales or Pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Long, H.J.; Yang, S.; Yin, X.H.; Zhao, Z.B.; Long, Y.H.; Fan, J.; Shu, R.; Gu, G.F. First Report of Leaf Spot on Prunus salicina Caused by Alternaria alternata in China. J. Plant Pathol. 2021, 103, 1021. [Google Scholar] [CrossRef]
- Al-Nadabi, H.H.; Maharachchikumbura, S.S.N.; Agrama, H.; Al-Azri, M.; Nasehi, A.; Al-Sadi, A.M. Molecular Characterization and Pathogenicity of Alternaria Species on Wheat and Date Palms in Oman. Eur. J. Plant Pathol. 2018, 152, 577–588. [Google Scholar] [CrossRef]
- Hahn, M.H.; Claus, A.; Koch, G.; Gelain, J.; Duarte, H.S.S. Occurrence of Alternaria arborescens Causing Leaf Spot on Pereskia aculeata in Brazil. Crop Prot. 2022, 151, 105824. [Google Scholar] [CrossRef]
- Achatz, G.; Oberkofler, H.; Lechenauer, E.; Simon, B.; Unger, A.; Kandler, D.; Ebner, C.; Prillinger, H.; Kraft, D.; Breitenbach, M. Molecular Cloning of Major and Minor Allergens of Alternaria alternata and Cladosporium herbarum. Mol. Immunol. 1995, 32, 213–227. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The Sections of Alternaria: Formalizing Species-Group Concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef]
- Morrison, V.A.; Weisdorf, D.J. Alternaria: A Sinonasal Pathogen of Immunocompromised Hosts. Clin. Infect. Dis. 1993, 16, 265–270. [Google Scholar] [CrossRef]
- Vartivarian, S.E.; Anaissie, E.J.; Bodey, G.P. Emerging Fungal Pathogens in Immunocompromised Patients: Classification, Diagnosis, and Management. Clin. Infect. Dis. 1993, 17, S487–S491. [Google Scholar] [CrossRef]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A Polyphasic Approach to Characterise Phoma and Related Pleosporalean Genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Woudenberg, J.H.C.; Cannon, P.F.; Crous, P.W. Colletotrichum Species with Curved Conidia from Herbaceous Hosts. Fungal Divers. 2009, 39, 45–87. [Google Scholar]
- Schubert, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; Starink, M.; Hill, C.F.; Zalar, P.; de Hoog, G.S.; Crous, P.W. Biodiversity in the Cladosporium herbarum Complex (Davidiellaceae, Capnodiales), with Standardisation of Methods for Cladosporium Taxonomy and Diagnostics. Stud. Mycol. 2007, 58, 105–156. [Google Scholar] [CrossRef]
- Hongsanan, S.; Jeewon, R.; Purahong, W.; Xie, N.; Liu, J.K.; Jayawardena, R.S.; Ekanayaka, A.H.; Dissanayake, A.; Raspé, O.; Hyde, K.D.; et al. Can We Use Environmental DNA as Holotypes? Fungal Divers. 2018, 92, 1–30. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous Identification of Fungi: Where Do We Stand and How Accurate and Precise Is Fungal DNA Barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Crous, P.W.; Rossman, A.Y.; Aime, M.C.; Allen, W.C.; Burgess, T.; Groenewald, J.Z.; Castlebury, L.A. Names of Phytopathogenic Fungi: A Practical Guide. Phytopathology 2021, 111, 1500–1508. [Google Scholar] [CrossRef]
- Stadler, M.; Lambert, C.; Wibberg, D.; Kalinowski, J.; Cox, R.J.; Kolařík, M.; Kuhnert, E. Intragenomic Polymorphisms in the ITS Region of High-Quality Genomes of the Hypoxylaceae (Xylariales, Ascomycota). Mycol. Prog. 2020, 19, 235–245. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Ryberg, M.; Kristiansson, E.; Abarenkov, K.; Larsson, K.-H.; Kõljalg, U. Taxonomic Reliability of DNA Sequences in Public Sequence Databases: A Fungal Perspective. PLoS ONE 2006, 1, e59. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Palm-Hernández, M.E. Systematics of Plant Pathogenic Fungi: Why It Matters. Plant Dis. 2008, 92, 1376–1386. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Wingfield, M.J.; Akulov, A.; Carnegie, A.J.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.Z.; Guarnaccia, V.; Halleen, F.; et al. Genera of Phytopathogenic Fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of Phytopathogenic Fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef] [PubMed]
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of Phytopathogenic Fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What Are Fungal Species and How to Delineate Them? Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Pinto, V.E.F.; Patriarca, A. Alternaria Species and Their Associated Mycotoxins. In Mycotoxigenic Fungi: Methods and Protocols, Methods in Molecular Biology; Moretti, A., Susca, A., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1542, pp. 13–32. ISBN 978-1-4939-6707-0. [Google Scholar]
- Kustrzeba-Wójcicka, I.; Siwak, E.; Terlecki, G.; Wolańczyk-Mędrala, A.; Mędrala, W. Alternaria alternata and Its Allergens: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2014, 47, 354–365. [Google Scholar] [CrossRef]
- Ghafri, A.A.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Al-Saady, N.A.; Al-Sadi, A.M. A New Section and a New Species of Alternaria Encountered from Oman. Phytotaxa 2019, 405, 279–289. [Google Scholar] [CrossRef]
- Gannibal, P.B.; Orina, A.S.; Gasich, E.L. A New Section for Alternaria Helianthiinficiens Found on Sunflower and New Asteraceous Hosts in Russia. Mycol. Prog. 2022, 21, 34. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria Themes and Variations (236-243) Host-Specific Toxin Producers. Mycotaxon 1999, 70, 325–369. [Google Scholar]
- Jayawardena, R.S.; Kandawatte, T.C.; Chethana, K.W.T.; Daranagama, D.A.; Dissanayake, A.J.; Goonasekara, I.D.; Manawasinghe; Mapook, A.; Jayasiri, S.C.; Karunarathna, A.; et al. Mycosphere Notes 102–168: Saprotrophic Fungi on Vitis in China, Italy, Russia and Thailand. Mycosphere 2018, 9, 1–114. [Google Scholar] [CrossRef]
- Simmons, E.G. Typification of Alternaria, Stemphylium, And Ulocladium. Mycologia 1967, 59, 67–92. [Google Scholar] [CrossRef]
- Kusaba, M.; Tsuge, T. Phylogeny of Alternaria Fungi Known to Produce Host-Specific Toxins on the Basis of Variation in Internal Transcribed Spacers of Ribosomal DNA. Curr. Genet. 1995, 28, 491–498. [Google Scholar] [CrossRef]
- Peever, T.L.; Su, G.; Carpenter-Boggs, L.; Timmer, L.W. Molecular Systematics of Citrus-Associated Alternaria Species. Mycologia 2004, 96, 119–134. [Google Scholar] [CrossRef]
- Armitage, A.D.; Cockerton, H.M.; Sreenivasaprasad, S.; Woodhall, J.; Lane, C.R.; Harrison, R.J.; Clarkson, J.P. Genomics Evolutionary History and Diagnostics of the Alternaria alternata Species Group Including Apple and Asian Pear Pathotypes. Front. Microbiol. 2020, 10, 3124. [Google Scholar] [CrossRef] [PubMed]
- Gannibal, P. Distribution of Alternaria Species among Sections. 2. Section Alternaria. Mycotaxon 2016, 130, 941–949. [Google Scholar] [CrossRef]
- Gannibal, P.; Lawrence, D. Distribution of Alternaria Species among Sections. 5. Species Producing Conidia with Many Longitudinal Septa. Mycotaxon 2018, 133, 285–291. [Google Scholar] [CrossRef]
- Wanasinghe, D.; Phukhamsakda, C.; Hyde, K.; Jeewon, R.; Lee, H.; Jones, E.; Tibpromma, S.; Tennakoon, D.; Dissanayake, A.; Jayasiri, S.; et al. Fungal Diversity Notes 709–839: Taxonomic and Phylogenetic Contributions to Fungal Taxa with an Emphasis on Fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Purahong, W.; Zhang, W.; Wubet, T.; Li, X.; Liu, M.; Zhao, W.S.; Hyde, K.D.; Liu, J.H.; Yan, J.Y. Biodiversity of Fungi on Vitis vinifera L. Revealed by Traditional and High-Resolution Culture-Independent Approaches. Fungal Divers. 2018, 90, 1–84. [Google Scholar] [CrossRef]
- Nishikawa, J.; Nakashima, C. Japanese Species of Alternaria and Their Species Boundaries Based on Host Range. Fungal Syst. Evol. 2020, 5, 197–282. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Piacentini, K.C.; Iwase, C.H.T.; Rocha, L. de O. Toxigenic Alternaria Species: Impact in Cereals Worldwide. Curr. Opin. Food Sci. 2018, 23, 57–63. [Google Scholar] [CrossRef]
- Kohmoto, K.; Itoh, Y.; Shimomura, N.; Kondoh, Y.; Otani, H.; Nishimura, S.; Nakatsuka, S. Isolation and Biological Activities of Two Host-Specific Toxins from the Tangerine Pathotype of Alternaria alternata. Phytopathology 1993, 83, 495–502. [Google Scholar] [CrossRef]
- Nishimura, S.; Kohmoto, K. Host-Specific Toxins and Chemical Structures from Alternaria Species. Annu. Rev. Phytopathol. 1983, 21, 87–116. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Sørensen, J.L.; Nielsen, K.F.; van den Ende, B.G.; de Hoog, S. A Polyphasic Approach to the Taxonomy of the Alternaria Infectoria Species–Group. Fungal Genet. Biol. 2009, 46, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Dongo, A.; Pryor, B.M. Secondary Metabolite Profiling of Alternaria dauci, A. porri, A. solani, and A. tomatophila. Mycol. Res. 2008, 112, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The Use of Secondary Metabolite Profiling in Chemotaxonomy of Filamentous Fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Aluthmuhandiram, J.V.S.; Chethana, K.W.T.; Manawasinghe, I.S.; Li, X.; Liu, M.; Hyde, K.D.; Phillips, A.J.L.; Zhang, W. Nigrospora Species Associated with Various Hosts from Shandong Peninsula, China. Mycobiology 2020, 48, 169–183. [Google Scholar] [CrossRef]
- Wang, M.; Liu, F.; Crous, P.W.; Cai, L. Phylogenetic Reassessment of Nigrospora: Ubiquitous Endophytes, Plant and Human Pathogens. Persoonia 2017, 39, 118–142. [Google Scholar] [CrossRef]
- Zhang, B. The Traits, Uses and Main Breeding Techniques of Celtis julianae. Shanghai Agric. Sci. Technol. 2014, 6, 104. [Google Scholar]
- Li, S.J.; Wu, Y.P.; Liu, Q.F. Effects of Phytohormones and Stratification Time on Germination of Celtis julianae Seeds. J. Northwest For. Univ. 2009, 24, 68–70. [Google Scholar]
- Zou, C.L.; Su, X.J.; Tong, A.; Yang, C.F.; Chen, H.Y.; Ji, W.L. Ornamental Characteristics and Ecological Effects of Celtis julianae. Mod. Hortic. 2022, 45, 26–29. [Google Scholar] [CrossRef]
- Xiao, Y.T.; Li, M.; Chen, F.M. Root Rot of Cinnamomum camphora (Linn) Presl Caused by Phytopythium vexans in China. Plants 2023, 12, 1072. [Google Scholar] [CrossRef]
- Ukwatta, K.M.; Lawrence, J.L.; Wijayarathna, C.D. The Study of Antimicrobial, Anti-Cancer, Anti-Inflammatory and α-Glucosidase Inhibitory Activities of Nigronapthaphenyl, Isolated from an Extract of Nigrospora sphaerica. Mycology 2019, 10, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Damm, U.; Mostert, L.; Crous, P.W.; Fourie, P.H. Novel Phaeoacremonium Species Associated with Necrotic Wood of Prunus Trees. Persoonia 2008, 20, 87–102. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Crous, P.W.; Schoch, C.L.; Hyde, K.D.; Wood, A.R.; Gueidan, C.; de Hoog, G.S.; Groenewald, J.Z. Phylogenetic Lineages in the Capnodiales. Stud. Mycol. 2009, 64, 17–47. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus Phylogenetics and the Origin of Known, Highly Virulent Pathogens, Inferred from ITS and Glyceraldehyde-3-Phosphate Dehydrogenase Gene Sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 Allergen Homologs from Alternaria and Related Taxa: Analysis of Phylogenetic Content and Secondary Structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; Spatafora, J.W. A Multi-Gene Phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of Localized Incongruence Using a Combinational Bootstrap Approach. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Symptoms of leaf blotch on Celtis julianae in the field. (A,B) Symptoms on diseased leaves; (C–E) diseased leaves on early stage, middle stage, and later stage, respectively; (F,G) conidial chains on a lesion of a leaf, scale bars: F = 100 μm, G = 50 μm.
Figure 1.
Symptoms of leaf blotch on Celtis julianae in the field. (A,B) Symptoms on diseased leaves; (C–E) diseased leaves on early stage, middle stage, and later stage, respectively; (F,G) conidial chains on a lesion of a leaf, scale bars: F = 100 μm, G = 50 μm.

Figure 2.
Pathogenicity of Alternaria isolates (B1, B2 and B3) on Celtis julianae. (A) No symptoms showing on the leaves from control plants 7 days after inoculation with sterile water. (B–D) Symptoms on leaves 7 days after inoculation with conidial suspensions of B1 (B), B2 (C), and B3 (D), respectively. Scale bars: (A–D) = 2 cm.
Figure 2.
Pathogenicity of Alternaria isolates (B1, B2 and B3) on Celtis julianae. (A) No symptoms showing on the leaves from control plants 7 days after inoculation with sterile water. (B–D) Symptoms on leaves 7 days after inoculation with conidial suspensions of B1 (B), B2 (C), and B3 (D), respectively. Scale bars: (A–D) = 2 cm.

Figure 3.
Phylogenetic relationship of Alternaria arborescens (B1, B2, B7, B8 and B9), and A. italica (B3, B4, B5 and B6) with related taxa derived from maximum-likelihood (ML) and Bayesian posterior probability analysis using concatenated ITS, LSU, SSU, GAPDH, Alt a 1, RPB2 and TEF1-α sequences of Alternaria spp., with Alternaria infectoria (CBS 210.86) as the outgroup. RA × ML bootstrap support values (ML ≥ 70) and Bayesian posterior probability values (PP ≥ 0.70) were shown at the nodes (ML/PP). Bar = 0.02 substitution per nucleotide position. The ex-type strains are in bold.
Figure 3.
Phylogenetic relationship of Alternaria arborescens (B1, B2, B7, B8 and B9), and A. italica (B3, B4, B5 and B6) with related taxa derived from maximum-likelihood (ML) and Bayesian posterior probability analysis using concatenated ITS, LSU, SSU, GAPDH, Alt a 1, RPB2 and TEF1-α sequences of Alternaria spp., with Alternaria infectoria (CBS 210.86) as the outgroup. RA × ML bootstrap support values (ML ≥ 70) and Bayesian posterior probability values (PP ≥ 0.70) were shown at the nodes (ML/PP). Bar = 0.02 substitution per nucleotide position. The ex-type strains are in bold.

Figure 4.
Morphological characteristics of Alternaria arborescens (isolate B1). (A) Front and (B) reverse view of 7-day-old colony on PCA; (C,D) conidiophores and conidia; (E) conidia; (F) conidial chains; (G–I) mycelia, conidiophores and conidial chains on PCA under a Zeiss stereo microscope.
Figure 4.
Morphological characteristics of Alternaria arborescens (isolate B1). (A) Front and (B) reverse view of 7-day-old colony on PCA; (C,D) conidiophores and conidia; (E) conidia; (F) conidial chains; (G–I) mycelia, conidiophores and conidial chains on PCA under a Zeiss stereo microscope.

Figure 5.
Morphological characteristics of Alternaria italica (isolate B3). (A) Front and (B) reverse view of 7-day-old colony on PCA. (C–E) Conidia, conidiophores and conidial chains. (F–H) Mycelia, conidiophores and conidial chains on PCA under a Zeiss stereo microscope.
Figure 5.
Morphological characteristics of Alternaria italica (isolate B3). (A) Front and (B) reverse view of 7-day-old colony on PCA. (C–E) Conidia, conidiophores and conidial chains. (F–H) Mycelia, conidiophores and conidial chains on PCA under a Zeiss stereo microscope.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fungi isolated from diseased leaves of Celtis julianae.
| Month | Number of Tissues | Number of Colonies | ||
|---|---|---|---|---|
| Nigrospora sp. | Alternaria sp. | Nothophoma sp. | ||
| September | 100 | 50 (63%) | 19 (24%) | 10 (13%) |
| October | 100 | 47 (54%) | 33 (38%) | 7 (8%) |
Table 2.
Primers used for PCR amplification in molecular identification of nine isolates (B1–B9).
| Locus | Primers | Primer Sequences (5′-3′) | Advantages and Limitations | Reference |
|---|---|---|---|---|
| ITS | ITS1 | TCCGTAGGTGAACCTGCGC | Universal fungal barcode, contains greater sequence variation, evolves faster, intragenomic variation gives the slow homogenization among the various copies | [64] |
| ITS4 | TCCTCCGCTTATTGATATGC | |||
| LSU | LROR | ACCCGCTGAACTTAAGC | Conserved and variable domain, low rate of molecular evolution reduces the taxonomic resolution at the species-level | [65] |
| LR5 | TCCTGAGGGAAACTTCG | |||
| SSU | NS1 | GTAGTCATATGCTTGTCTC | [64] | |
| NS4 | CTTCCGTCAATTCCTTTAAG | |||
| GAPDH | GPD1 | CAACGGCTTCGGTCGCATTG | Highly effective for heterologous protein expression in microorganisms, the expression level may increase under inducing treatments | [66] |
| GPD2 | GCCAAGCAGTTGGTTGTGC | |||
| Alt a 1 | Alt-al-for | ATGCAGTTCACCACCATCGC | A gene for the Alternaria major allergen, supports grouping of Alternaria spp. and related taxa | [67] |
| Alt-a1-rev | ACGAGGGTGAYGTAGGCGTC | |||
| RPB2 | RPB2-5F2 | GGGGWGAYCAGAAGAAGGC | Recover well-supported clades at shallow and deep taxonomic levels and has a better species-resolving power than rDNA markers | [68] |
| RPB2-7cR | CCCATRGCT TGT YYRCCCAT | |||
| TEF1-α | EF1-728F | CATCGAGAAGTTCGAGAAGG | Recover some deep and ordinal-level relationships but with greater branch support from nucleotides | [69] |
| EF1-986R | TACTTGAAGGAACCCTTACC |
Table 3.
Isolates of Alternaria spp. used in this study and corresponding GenBank accession numbers.
Table 3.
Isolates of Alternaria spp. used in this study and corresponding GenBank accession numbers.
| Species | Isolate | Locality, Host/Substrate | Accession Numbers | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | LSU | SSU | GAPDH | Alt a 1 | RPB2 | TEF1-α | |||
| Alternaria alstroemeriae | CBS 118808 | USA, Alstroemeria sp. | KP124296 | KP124447 | KP124917 | KP124153 | KP123845 | KP124764 | KP125071 |
| A. alternantherae | CBS 124392 | China, Solanum melongena | KC584179 | KC584251 | KC584506 | KC584096 | KP123846 | KC584374 | KC584633 |
| A. alternata | CBS 916.96 | India, Arachis hypogaea | AF347031 | DQ678082 | KC584507 | AY278808 | AY563301 | KC584375 | KC584634 |
| A. alternata | CBS 112249 | -, - | KP124338 | KP124490 | KP124960 | KP124192 | KP123886 | KP124806 | KP125114 |
| A. arborescens | CBS 102605 | USA, Solanum lycopersicum | AF347033 | KC584253 | KC584509 | AY278810 | AY563303 | KC584377 | KC584636 |
| A. arborescens | B1 = CFCC 59038 * | China, Celtis julianae | OQ691659 | OQ692430 | OQ692438 | OQ710109 | OQ710107 | OQ710111 | OQ710113 |
| A. arborescens | B2 = CFCC 59039 * | China, Celtis julianae | OQ691640 | OQ692431 | OQ692439 | OQ710110 | OQ710108 | OQ710112 | OQ710114 |
| A. arborescens | B7 * | China, Celtis julianae | OR243734 | OR366490 | OR366484 | OR475216 | OR475232 | OR475224 | OR475210 |
| A. arborescens | B8 * | China, Celtis julianae | OR243735 | OR366491 | OR366485 | OR475217 | OR475231 | OR475225 | OR475211 |
| A. arborescens | B9 * | China, Celtis julianae | OR243736 | OR366492 | OR366486 | OR475218 | OR475233 | OR475226 | OR475212 |
| A. arborescens | CBS 119545 | New Zealand, Senecio skirrhodon | KP124409 | KP124562 | KP125032 | KP124260 | KP123956 | KP124879 | KP125187 |
| A. arctoseptata | MFLUCC 21-0139 | Italy, Lathyrus sp. (Fabaceae) | - | MZ621948 | MZ621874 | 0K236608 | OK236755 | OK236655 | OK236702 |
| A. avenicola | CBS 121459 | Norway, Avena sp. | KC584183 | KC584256 | KC584512 | KC584100 | - | KC584380 | KC584639 |
| A. axiaeriisporifera | CBS 118715 | New Zealand, Gypsophila paniculata | KC584184 | KC584257 | KC584513 | KC584101 | - | KC584381 | KC584640 |
| A. baoshanensis | MFLUCC 21-0124 | China, Curcubita moschata | MZ622003 | MZ621952 | MZ621878 | OK236613 | OK236760 | OK236659 | OK236706 |
| A. betae-kenyensis | CBS 118810 | Kenya, Beta vulgaris var. cicla | KP124419 | KP124572 | KP125042 | KP124270 | KP123966 | KP124888 | KP125197 |
| A. brassicae | CBS 116528 | USA, Brassica oleracea | KC584185 | KC584258 | KC584514 | KC584102 | - | KC584382 | KC584641 |
| A. brassicicola | CBS 118699 | USA, Brassica oleracea | JX499031 | KC584259 | KC584515 | KC584103 | - | KC584383 | KC584642 |
| A. breviconidiophora | MFLUCC 22-0075 | Italy, Digitalis sp. (Scrophulariaceae) | MZ621997 | MZ621944 | MZ621870 | OK236604 | OK236751 | OK236651 | OK236698 |
| A. burnsii | CBS 107.38 | India, Cuminum cyminum | KP124420 | KP124573 | KP125043 | JQ646305 | KP123967 | KP124889 | KP125198 |
| A. carotiincultae | CBS 109381 | USA, Daucus carota | KC584188 | KC584262 | KC584518 | KC584106 | - | KC584386 | KC584645 |
| A. cinerariae | CBS 116495 | USA, Ligularia sp. | KC584190 | KC584265 | KC584521 | KC584109 | - | KC584389 | KC584648 |
| A. conoidea | CBS 132.89 | Saudi Arabia, Ricinus communis | FJ348226 | KC584327 | KC584585 | FJ348227 | FJ348228 | KC584452 | KC584711 |
| A. dauci | CBS 117097 | USA, Daucus carota | KC584192 | KC584268 | KC584524 | KC584111 | KJ718678 | KC584392 | KC584651 |
| A. doliconidium | KUN-HKAS 100840T | Italy, Rosa canina | NR158361 | NG069551 | NG065142 | - | - | - | - |
| A. eichhorniae | CBS 489.92 | India, Eichhornia crassipes | KC146356 | KP124579 | KP125049 | KP124276 | KP123973 | KP124895 | KP125204 |
| A. ellipsoidea | CBS 119674 | USA, Dianthus barbatus | KC584196 | KC584272 | KC584528 | KC584115 | - | KC584396 | KC584655 |
| A. ellipsoidialis | MFLUCC 21-0132 | Italy, Brassica sp. (Brassicaceae) | MZ621989 | MZ621936 | MZ621862 | OK236596 | OK236743 | OK236643 | OK236690 |
| A. eupatoriicola | MFLUCC 21-0122 | Italy, Eupatorium cannabinum (Asteraceae) | MZ621982 | MZ621929 | MZ621855 | OK236589 | OK236736 | OK236636 | OK236683 |
| A. falcata | MFLUCC 21-0123 | Italy, Atriplex sp. (Chenopodiaceae) | MZ621992 | MZ62139 | MZ621865 | OK236599 | OK236746 | OK236649 | OK236693 |
| A. gaisen | CBS 118488 | Japan, Pyrus pyrifolia | KP124427 | KP124581 | KP125051 | KP124278 | KP123975 | KP124897 | KP125206 |
| A. gossypina | CBS 104.32 | Zimbabwe, Gossypium sp. | KP124430 | KP124584 | KP125054 | JQ646312 | JQ646395 | KP124900 | KP125209 |
| A. gypsophilae | CBS 107.41 | Netherlands, Gypsophila elegans | KC584199 | KC584277 | KC584533 | KC584118 | KJ718688 | KC584401 | KC584660 |
| A. helianthiinficiens | CBS 117370 | UK, Helianthus annuus | KC584200 | KC584278 | KC584534 | KC584119 | - | KC584402 | KC584661 |
| A. helianthiinficiens | CBS 208.86 | USA, Helianthus annuus | JX101649 | KC584279 | KC584535 | KC584120 | - | KC584403 | EU130548 |
| A. infectoria | CBS 210.86 | UK, Triticum aestivum | DQ323697 | KC584280 | KC584536 | AY278793 | FJ266502 | KC584404 | KC584662 |
| A. iridiaustralis | CBS 118486 | Australia, Iris sp. | KP124435 | KP124589 | KP125059 | KP124284 | KP123981 | KP124905 | KP125214 |
| A. italica | MFLUCC 14-0421T | Italy, Pleosporaceae | MG764017 | MG818319 | - | - | - | MG859737 | - |
| A. italica | B3 = CFCC 59359 * | China, Celtis julianae | OR272062 | OR366487 | OR366480 | OR475213 | OR475227 | OR475220 | OR475207 |
| A. italica | B4 = CFCC 59309 * | China, Celtis julianae | OR243731 | OR366526 | OR366481 | OR475219 | OR475228 | OR475221 | OR250485 |
| A. italica | B5 = CFCC 59310 * | China, Celtis julianae | OR243732 | OR366488 | OR366482 | OR475214 | OR475229 | OR475222 | OR475208 |
| A. italica | B6 = CFCC 59311 * | China, Celtis julianae | OR243733 | OR366489 | OR366483 | OR475215 | OR475230 | OR475223 | OR475209 |
| A. jacinthicola | CBS 133751 | Mali, Eichhornia crassipes | KP124438 | KP124592 | KP125062 | KP124287 | KP123984 | KP124908 | KP125217 |
| A. juxtiseptata | CBS 119673 | Australia, Gypsophila paniculata | KC584202 | KC584282 | KC584538 | KC584122 | - | KC584406 | KC584664 |
| A. lathyri | MFLUCC 21-0140 | Italy, Lathyrus sp. (Fabaceae) | MZ621974 | MZ621921 | MZ621847 | OK236581 | OK236728 | OK236628 | OK236675 |
| A. longipes | CBS 540.94 | USA, Nicotiana tabacum | AY278835 | KC584285 | KC584541 | AY278811 | AY563304 | KC584409 | KC584667 |
| A. macroconidia | MFLUCC 21-0134 | Italy, Spartium junceum | MZ622001 | MZ621950 | MZ621876 | OK236610 | OK236757 | OK236657 | OK236704 |
| A. macrospora | CBS 117228 | USA, Gossypium barbadense | KC584204 | KC584286 | KC584542 | KC584124 | KJ718702 | KC584410 | KC584668 |
| A. mimicula | CBS 118696 | USA, Lycopersicon esculentum | FJ266477 | KC584287 | KC584543 | AY562415 | GQ180094 | KC584411 | KC584669 |
| A. minimispora | MFLUCC 21-0127 | Thailand, Citrullus lanatus | MZ621980 | MZ621927 | MZ621853 | OK236587 | OK236734 | OK236634 | OK236681 |
| A. muriformispora | MFLUCC 22-0073 | Italy, Plantago sp. (Plantaginaceae) | MZ621976 | MZ621923 | MZ621849 | OK236583 | OK236730 | OK236630 | OK236677 |
| A. nobilis | CBS 116490 | New Zealand, Dianthus caryophyllus | KC584208 | KC584291 | KC584547 | KC584127 | - | KC584415 | KC584673 |
| A. oblongoellipsoidea | MFLUCC 22-0074 | Italy, Cichorium intybus | MZ621967 | MZ621914 | MZ621840 | OK236574 | OK236721 | OK236621 | OK236668 |
| A. obpyriconidia | MFLUCC 21-0121 | Italy, Vicia faba | MZ621978 | MZ621925 | MZ621851 | OK236585 | OK236732 | OK236633 | OK236680 |
| A. orobanches | MFLUCC 21-0137 | Italy, Orobanche sp. | MZ622007 | MZ621956 | MZ621882 | - | OK236763 | - | OK236710 |
| A. ovoidea | MFLUCC 14-0427 | Italy, Dactylis glomerata | MZ622005 | MZ621954 | MZ621880 | OK236614 | OK236761 | OK236661 | OK236708 |
| A. perpunctulata | CBS 115267 | USA, Alternanthera philoxeroides | KC584210 | KC584294 | KC584550 | KC584129 | JQ905111 | KC584418 | KC584676 |
| A. petroselini | CBS 112.41 | –, Petroselinum sativum | KC584211 | KC584295 | KC584551 | KC584130 | - | KC584419 | KC584677 |
| A. phragmiticola | MFLUCC 21-0125 | Italy, Phragmites sp. | MZ621994 | MZ621941 | MZ621867 | OK236602 | OK236749 | OK236649 | OK236696 |
| A. porri | CBS 116698 | USA, Allium cepa | DQ323700 | KC584297 | KC584553 | KC584132 | KJ718726 | KC584421 | KC584679 |
| A. pseudoinfectoria | MFLUCC 21-0126 | Italy, Chenopodium sp. | MZ621984 | MZ621931 | MZ621857 | OK236591 | OK236738 | OK236638 | OK236685 |
| A. pseudorostrata | CBS 119411 | USA, Euphorbia pulcherrima | JN383483 | KC584298 | KC584554 | AY562406 | AY563295 | KC584422 | KC584680 |
| A. radicina | CBS 245.67 | USA, Daucus carota | KC584213 | KC584299 | KC584555 | KC584133 | FN689405 | KC584423 | KC584681 |
| A. rostroconidia | MFLUCC 21-0136 | Italy, Arabis sp. | MZ621969 | MZ621916 | MZ621842 | OK236576 | OK236723 | OK236623 | OK236670 |
| A. salicicola | MFLUCC 22-0072 | Russia, Salix alba | MZ621999 | MZ621946 | MZ621872 | OK236606 | OK236753 | OK236653 | OK236700 |
| A. saponariae | CBS 116492 | USA, Saponaria officinalis | KC584215 | KC584301 | KC584557 | KC584135 | - | KC584425 | KC584683 |
| A. selini | CBS 109382 | Saudi Arabia, Petroselinum crispum | AF229455 | KC584302 | KC584558 | AY278800 | FJ266504 | KC584426 | KC584684 |
| A. septorioides | CBS 106.41 | Netherlands, Reseda odorata | KC584216 | KC584303 | KC584559 | KC584136 | - | KC584427 | KC584685 |
| A. setosa | YZU 191101 | China, Iris japonica | OP2341770 | - | - | OP352306 | OP352294 | OP352294 | OP374459 |
| A. smyrnii | CBS 109380 | UK, Smyrnium olusatrum | AF229456 | KC584305 | KC584561 | KC584138 | - | KC584429 | KC584687 |
| A. solani | CBS 116651 | USA, Solanum tuberosum | KC584217 | KC584306 | KC584562 | KC584139 | GQ180097 | KC584430 | KC584688 |
| A. solidaccana | CBS 118698 | Bangladesh, Soil | KC584219 | KC584308 | KC584564 | KC584141 | - | KC584432 | KC584690 |
| A. sonchi | CBS 119675 | Canada, Sonchus asper | KC584220 | KC584309 | KC584565 | KC584142 | - | KC584433 | KC584691 |
| A. tectorum | YZU 161050 | China, Iris tectorum | OP341728 | - | - | OP352303 | OP293714 | OP352291 | OP374456 |
| A. tectorum | YZU 161052 | China, Iris tectorum | - | - | OP341817.1 | OP352304.1 | OP293715.1 | OP352292.1 | OP374457.1 |
| A. tomato | CBS 103.30 | Unknown, Solanum lycopersicum | KP124445 | KP124599 | KP125069 | KP124294 | KP123991 | KP124915 | KP125224 |
| A. torilis | MFLUCC 14-0433 | Italy, Torilis arvensis | MZ621988 | MZ621935 | MZ621861 | OK236594 | OK236741 | OK236641 | OK236688 |
| A. vaccariae | CBS 116533 | USA, Vaccaria hispanica | KC584223 | KC584314 | KC584570 | KC584146 | JQ646386 | KC584438 | KC584696 |
| A. vaccariicola | CBS 118714 | USA, Vaccaria hispanica | KC584224 | KC584315 | KC584571 | KC584147 | JQ646384 | KC584439 | KC584697 |
| A. vitis | MFLUCC 17-1109T | Chile, China, El Salvador, Greece, India, Italy, Romania, Russia, Thailand, Turkmenistan, Pleosporaceae | MG764007 | - | - | - | - | - | - |
* Isolates in this study. Ex-type cultures are shown in bold. Abbreviations: CBS: Culture collection of the Westerdijk Fungal Biodiversity Institute, Utrecht, The Netherlands; CFCC: China Forestry Culture Collection Center, Beijing, China; KUN-HKAS: Herbarium of Cryptogams Kunming Institute of Botany Academia Sinica, Yunnan, China; MFLUCC: Mae Fah Luang University Culture Collection, Chiang Rai, Thailand; YZU: Fungal Herbarium of Yangtze University, Jingzhou, Hubei, China.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liao, Y.-C.-Z.; Cao, Y.-J.; Wan, Y.; Li, H.; Li, D.-W.; Zhu, L.-H. Alternaria arborescens and A. italica Causing Leaf Blotch on Celtis julianae in China. Plants 2023, 12, 3113. https://doi.org/10.3390/plants12173113
AMA Style
Liao Y-C-Z, Cao Y-J, Wan Y, Li H, Li D-W, Zhu L-H. Alternaria arborescens and A. italica Causing Leaf Blotch on Celtis julianae in China. Plants. 2023; 12(17):3113. https://doi.org/10.3390/plants12173113
Chicago/Turabian StyleLiao, Yang-Chun-Zi, Yi-Jia Cao, Yu Wan, Hui Li, De-Wei Li, and Li-Hua Zhu. 2023. "Alternaria arborescens and A. italica Causing Leaf Blotch on Celtis julianae in China" Plants 12, no. 17: 3113. https://doi.org/10.3390/plants12173113
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.