
Diterpene Resin Acids and Olefins in Calabrian Pine (Pinus nigra subsp. laricio (Poiret) Maire) Oleoresin: GC-MS Profiling of Major Diterpenoids in Different Plant Organs, Molecular Identification and Expression Analysis of Diterpene Synthase Genes

 ,
,  ,
,  , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. In the Pinaceae, the Diterpene Metabolites Profiles Are Tissue-Specific and Species-Specific
2.2. A Phylogeny-Based Approach for Isolating Partial and Full-Length cDNAs Coding for Diterpene Synthases in Calabrian Pine
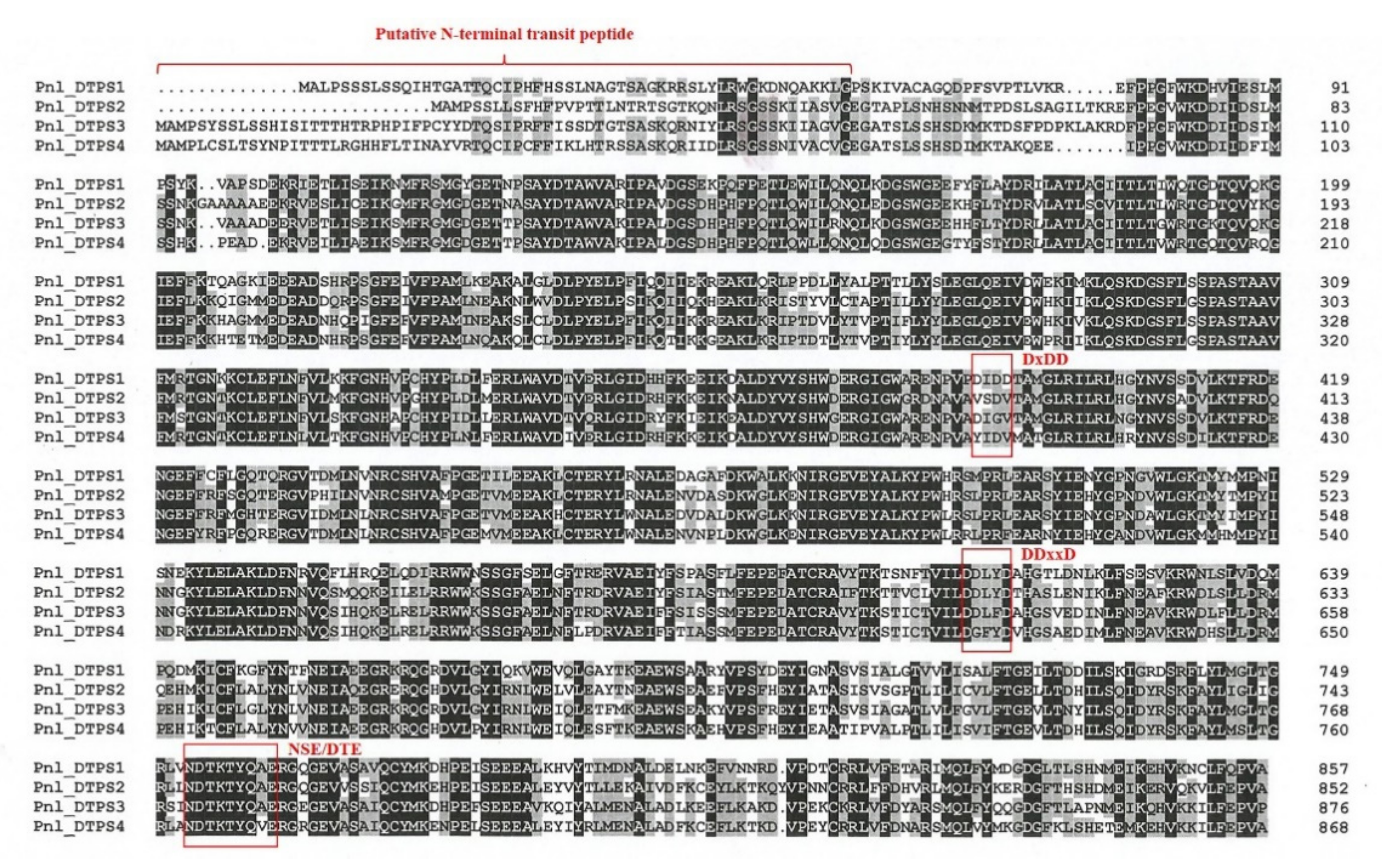
2.3. Sequence-Based Analysis Predicts That Both Monofunctional and Bifunctional Diterpene Synthases Are Involved in the Biosynthesis of Diterpene Resin Acids in Calabrian Pine
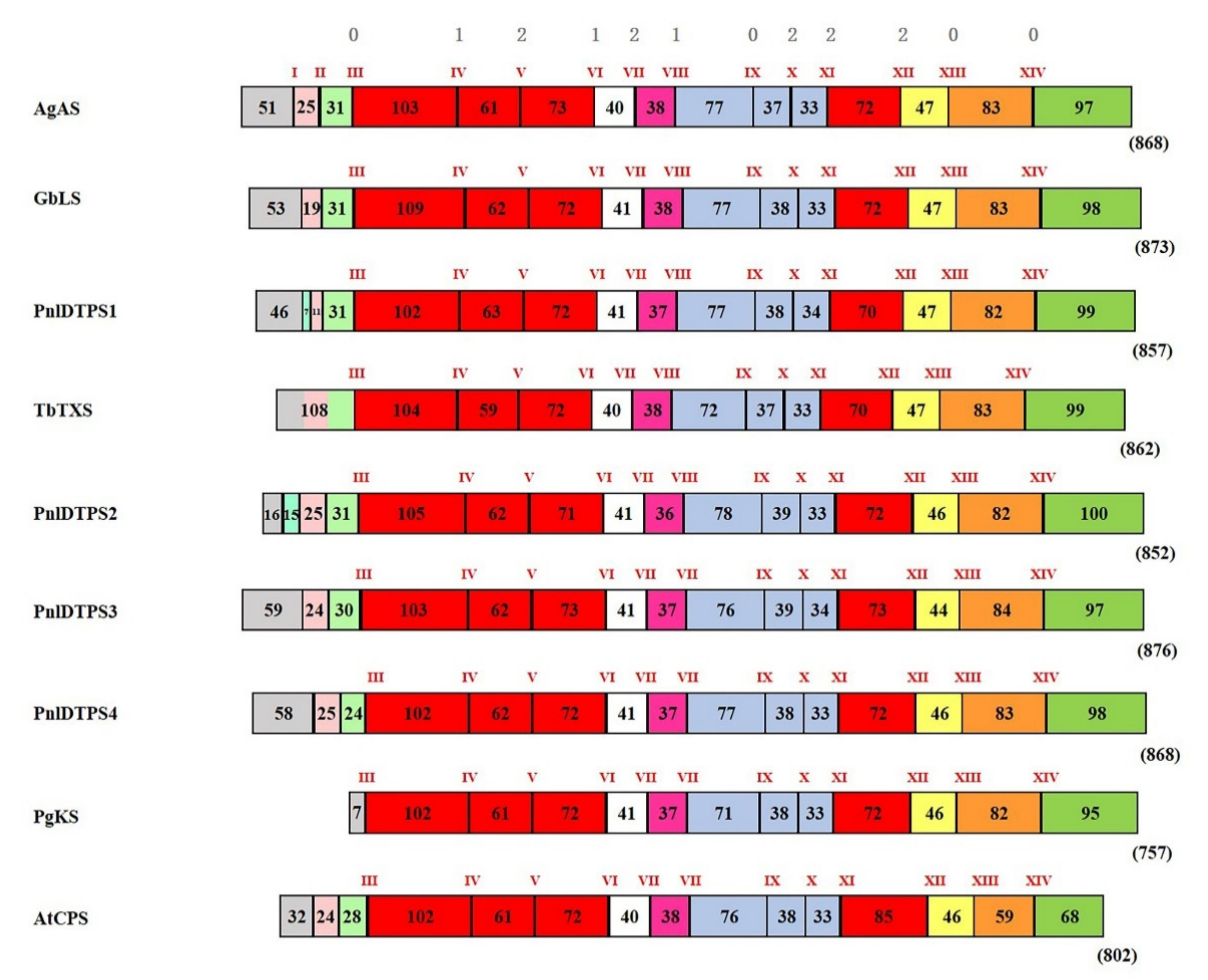
2.4. Genomic Organization of Diterpene Synthases in Calabrian Pine on the Background of DTPS Functional Evolution
2.5. Transcripts Profiling of Calabrian Pine DTPS Genes Reveal Differential Expression across Different Tissues and Suggest Their Putative Roles in the Biosynthesis of Diterpene Resin Acids
3. Materials and Methods
3.1. Plant Material
3.2. Extraction and GC/MS Analysis of Diterpene Metabolites
3.3. RNA Isolation and cDNA Synthesis
3.4. DNA Extraction
3.5. Isolation of Partial and Full-Length cDNAs Coding for Diterpene Synthases
3.6. Isolation of Genomic Sequences Coding for Diterpene Synthases
3.7. Cloning and Sequencing of RACE, cDNA and Genomic Amplification Products
3.8. Analysis of the Nucleotide and of the Deduced Amino Acid Sequences
3.9. Phylogenetic Analysis
3.10. Gene Expression Analysis
3.11. Statistical Analysis
3.12. Ethical Statement
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zulak, K.G.; Bohlmann, J. Terpenoid biosynthesis and specialized vascular cells of conifer defense. J. Integr. Plant Biol. 2010, 52, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Celedon, J.M.; Bohlmann, J. Oleoresin defenses in conifers: Chemical diversity, terpene synthases and limitations of oleoresin defense under climate change. New Phytol. 2019, 224, 1444–1463. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Lee, Y.; Tsao, N.; Wang, S.; Zerbe, P.; Chu, F. Biochemical characterization of diterpene synthases of Taiwania cryptomerioides expands the known functional space of specialized diterpene metabolism in gymnosperms. Plant J. 2019, 100, 1254–1272. [Google Scholar] [CrossRef]
- Boncan, D.A.T.; Tsang, S.S.; Li, C.; Lee, I.H.; Lam, H.-M.; Chan, T.-F.; Hui, J.H. Terpenes and terpenoids in plants: Interactions with environment and insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef]
- Chou, Y.-W.; Thomas, P.I.; Ge, X.-J.; LePage, B.A.; Wang, C.-N. Refugia and phylogeography of Taiwania in East Asia. J. Biogeogr. 2011, 38, 1992–2005. [Google Scholar] [CrossRef]
- Leslie, A.B.; Beaulieu, J.M.; Rai, H.S.; Crane, P.; Donoghue, M.J.; Mathews, S. Hemisphere-scale differences in conifer evolutionary dynamics. Proc. Natl. Acad. Sci. USA 2012, 109, 16217–16221. [Google Scholar] [CrossRef] [Green Version]
- Karchesy, J.J.; Kelsey, R.G.; González-Hernández, M.P. Yellow-cedar, Callitropsis (Chamaecyparis) nootkatensis, secondary metabolites, biological activities, and chemical ecology. J. Chem. Ecol. 2018, 44, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Simoneit, B.R.T.; Cox, R.E.; Oros, D.R.; Otto, A. Terpenoid compositions of resins from Callitris species (Cupressaceae). Molecules 2018, 23, 3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, J.A.; Madilao, L.L.; White, R.; Yanchuk, A.; King, J.; Bohlmann, J. Terpenoid metabolite profiling in Sitka spruce identifies association of dehydroabietic acid, (+)-3-carene, and terpinolene with resistance against white pine weevil. Botany 2010, 88, 810–820. [Google Scholar] [CrossRef]
- Lah, L.; Haridas, S.; Bohlmann, J.; Breuil, C. The cytochromes P450 of Grosmannia clavigera: Genome organization, phylogeny, and expression in response to pine host chemicals. Fungal Genet. Biol. 2013, 50, 72–81. [Google Scholar] [CrossRef]
- Wang, Y.; Lim, L.; DiGuistini, S.; Robertson, G.; Bohlmann, J.; Breuil, C. A specialized ABC efflux transporter G c ABC-G 1 confers monoterpene resistance to Grosmannia clavigera, a bark beetle-associated fungal pathogen of pine trees. New Phytol. 2013, 197, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Kshatriya, K.; Whitehill, J.G.; Madilao, L.; Henderson, H.; Kermode, A.; Kolotelo, D.; Bohlmann, J.; Kolotelo, D. Histology of resin vesicles and oleoresin terpene composition of conifer seeds. Can. J. For. Res. 2018, 48, 1073–1084. [Google Scholar] [CrossRef]
- Zerbe, P.; Hamberger, B.; Yuen, M.M.; Chiang, A.; Sandhu, H.K.; Madilao, L.L.; Nguyen, A.; Hamberger, B.; Bach, S.S.; Bohlmann, J. Gene discovery of modular diterpene metabolism in nonmodel systems. Plant Physiol. 2013, 162, 1073–1091. [Google Scholar] [CrossRef] [Green Version]
- Mafu, S.; Karunanithi, P.S.; Palazzo, T.A.; Harrod, B.L.; Rodriguez, S.M.; Mollhoff, I.N.; O’Brien, T.E.; Tong, S.; Fiehn, O.; Tantillo, D.J.; et al. Biosynthesis of the microtubule-destabilizing diterpene pseudolaric acid B from golden larch involves an unusual diterpene synthase. Proc. Natl. Acad. Sci. USA 2017, 114, 974–979. [Google Scholar] [CrossRef] [Green Version]
- Croteau, R.; Ketchum, R.E.B.; Long, R.M.; Kaspera, R.; Wildung, M.R. Taxol biosynthesis and molecular genetics. Phytochem. Rev. 2006, 5, 75–97. [Google Scholar] [CrossRef] [Green Version]
- Zerbe, P.; Chiang, A.; Yuen, M.; Hamberger, B.; Hamberger, B.; Draper, J.A.; Britton, R.; Bohlmann, J. Bifunctional cis-abienol synthase from Abies balsamea discovered by transcriptome sequencing and its implications for diterpenoid fragrance production. J. Biol. Chem. 2012, 287, 12121–12131. [Google Scholar] [CrossRef] [Green Version]
- Hamberger, B.; Ohnishi, T.; Hamberger, B.; Séguin, A.; Bohlmann, J. Evolution of diterpene metabolism: Sitka spruce CYP720B4 catalyzes multiple oxidations in resin acid biosynthesis of conifer defense against insects. Plant Physiol. 2011, 157, 1677–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbe, P.; Bohlmann, J. Enzymes for Synthetic Biology of Ambroxide-Related Diterpenoid Fragrance Compounds. In Signaling Pathways for Translation; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2015; Volume 148, pp. 427–447. [Google Scholar]
- Gao, Y.; Honzatko, R.B.; Peters, R.J. Terpenoid synthase structures: A so far incomplete view of complex catalysis. Nat. Prod. Rep. 2012, 29, 1153–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicandri, E.; Paolacci, A.R.; Osadolor, S.; Sorgonà, A.; Badiani, M.; Ciaffi, M. On the evolution and functional diversity of terpene synthases in the Pinus species: A review. J. Mol. Evol. 2020, 88, 253–283. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.J. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar] [CrossRef] [Green Version]
- Hall, D.E.; Zerbe, P.; Jancsik, S.; Quesada, A.L.; Dullat, H.; Madilao, L.L.; Yuen, M.; Bohlmann, J. Evolution of conifer diterpene synthases: Diterpene resin acid biosynthesis in lodgepole pine and jack pine involves monofunctional and bifunctional diterpene synthases. Plant Physiol. 2013, 161, 600–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.-T.; Wang, C.-H.; Hon, C.-Y.; Lee, Y.-R.; Chu, F.-H. Discovery and characterization of diterpene synthases in Chamaecyparis formosensis Matsum. which participated in an unprecedented diterpenoid biosynthesis route in conifer. Plant Sci. 2021, 304, 110790. [Google Scholar] [CrossRef] [PubMed]
- Keeling, C.I.; Weisshaar, S.; Ralph, S.G.; Jancsik, S.; Hamberger, B.; Dullat, H.K.; Bohlmann, J. Transcriptome mining, functional characterization, and phylogeny of a large terpene synthase gene family in spruce (Picea spp.). BMC Plant Biol. 2011, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Foti, V.; Araniti, F.; Manti, F.; Alicandri, E.; Giuffrè, A.M.; Bonsignore, C.P.; Castiglione, E.; Sorgonà, A.; Covino, S.; Paolacci, A.R.; et al. Profiling volatile terpenoids from calabrian pine stands infested by the pine processionary moth. Plants 2020, 9, 1362. [Google Scholar] [CrossRef] [PubMed]
- Nicolaci, A.; Travaglini, D.; Menguzzato, G.; Nocentini, S.; Veltri, A.; Iovino, F. Ecological and anthropogenic drivers of Calabrian pine (Pinus nigra J.F. Arn. ssp. laricio (Poiret) Maire) distribution in the Sila mountain range. iForest—Biogeosci. For. 2015, 8, 497–508. [Google Scholar] [CrossRef] [Green Version]
- López-Goldar, X.; Lundborg, L.; Borg-Karlson, A.K.; Zas, R.; Sampedro, L. Resin acids as inducible chemical defences of pine seedlings against chewing insects. PLoS ONE 2020, 15, e0232692. [Google Scholar] [CrossRef]
- Vogel, B.S.; Wildung, M.R.; Vogel, G.; Croteau, R. Abietadiene synthase from grand fir (Abies grandis) cDNA isolation, characterization, and bacterial expression of a bifunctional diterpene cyclase involved in resin acid biosynthesis. J. Biol. Chem. 1996, 271, 23262–23268. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.M.; Faldt, J.; Bohlmann, J. Functional Characterization of nine Norway spruce TPS genes and evolution of gymnosperm terpene synthases of the TPS-d subfamily. Plant Physiol. 2004, 135, 1908–1927. [Google Scholar] [CrossRef] [Green Version]
- Ro, D.-K.; Bohlmann, J. Diterpene resin acid biosynthesis in loblolly pine (Pinus taeda): Functional characterization of abietadiene/levopimaradiene synthase (PtTPS-LAS) cDNA and subcellular targeting of PtTPS-LAS and abietadienol/abietadienal oxidase (PtAO, CYP720B1). Phytochemistry 2006, 67, 1572–1578. [Google Scholar] [CrossRef]
- Peters, R.J.; Croteau, R.B. Abietadiene synthase catalysis: Conserved residues involved in protonation-initiated cyclization of geranylgeranyl diphosphate to (+)-copalyl diphosphate. Biochemistry 2002, 41, 1836–1842. [Google Scholar] [CrossRef]
- Köksal, M.; Hu, H.; Coates, R.M.; Peters, R.J.; Christianson, D.W. Structure and mechanism of the diterpene cyclase ent-copalyl diphosphate synthase. Nat. Chem. Biol. 2011, 7, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.E.; Yuen, M.M.S.; Jancsik, S.; Quesada, A.L.; Dullat, H.K.; Li, M.; Henderson, H.; Arango-Velez, A.; Liao, N.Y.; Docking, R.T.; et al. Transcriptome resources and functional characterization of monoterpene synthases for two host species of the mountain pine beetle, lodgepole pine (Pinus contorta) and jack pine (Pinus banksiana). BMC Plant Biol. 2013, 13, 80. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.W.S.; Simpson, C.G. Splice site selection in plant pre-mRNA splicing. Annu. Rev. Plant Biol. 1998, 49, 77–95. [Google Scholar] [CrossRef]
- Li, W. Molecular Evolution; Sinauer Associates Incorporated: Sunderland, MA, USA, 1997. [Google Scholar]
- Trapp, S.C.; Croteau, R.B. Genomic Organization of plant terpene synthases and molecular evolutionary implications. Genetics 2001, 158, 811–832. [Google Scholar] [CrossRef] [PubMed]
- Keeling, C.I.; Dullat, H.K.; Yuen, M.; Ralph, S.G.; Jancsik, S.; Bohlmann, J. Identification and functional characterization of monofunctionalent-copalyl diphosphate and ent-kaurene synthases in white spruce reveal different patterns for diterpene synthase evolution for primary and secondary metabolism in gymnosperms. Plant Physiol. 2009, 152, 1197–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Pavy, N.; Boyle, B.; Nelson, C.; Paule, C.; Giguère, I.; Caron, S.; Parsons, L.; Dallaire, N.; Bedon, F.; Bérubé, H.; et al. Identification of conserved core xylem gene sets: Conifer cDNA microarray development, transcript profiling and computational analyses. New Phytol. 2008, 180, 766–786. [Google Scholar] [CrossRef] [PubMed]
- Hebsgaard, S.M. Splice site prediction in Arabidopsis thaliana pre-mRNA by combining local and global sequence information. Nucleic Acids Res. 1996, 24, 3439–3452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emanuelsson, O.; Nielsen, H.; Von Heijne, G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. 1999, 8, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Small, I.; Peeters, N.; Legeai, F.; Lurin, C. Predotar: A tool for rapidly screening proteomes for N-terminal targeting sequences. Proteomics 2004, 4, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Mega, X. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, H.; Yang, Z.; Hu, Y.; Tan, J.; Jia, J.; Xu, H.; Chen, X. Reference genes selection for quantitative gene expression studies in Pinus massoniana L. Trees 2016, 30, 685–696. [Google Scholar] [CrossRef]
- Mo, J.; Xu, J.; Jin, W.; Yang, L.; Yin, T.; Shi, J. Identification of reference genes for quantitative gene expression studies in Pinus massoniana and its introgression hybrid. Forests 2019, 10, 787. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Paolacci, A.R.; Catarcione, G.; Ederli, L.; Zadra, C.; Pasqualini, S.; Badiani, M.; Musetti, R.; Santi, S.; Ciaffi, M. Jasmonate-mediated defence responses, unlike salicylate-mediated responses, are involved in the recovery of grapevine from bois noir disease. BMC Plant Biol. 2017, 17, 118. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alicandri, E.; Covino, S.; Sebastiani, B.; Paolacci, A.R.; Badiani, M.; Manti, F.; Bonsignore, C.P.; Sorgonà, A.; Ciaffi, M. Diterpene Resin Acids and Olefins in Calabrian Pine (Pinus nigra subsp. laricio (Poiret) Maire) Oleoresin: GC-MS Profiling of Major Diterpenoids in Different Plant Organs, Molecular Identification and Expression Analysis of Diterpene Synthase Genes. Plants 2021, 10, 2391. https://doi.org/10.3390/plants10112391
Alicandri E, Covino S, Sebastiani B, Paolacci AR, Badiani M, Manti F, Bonsignore CP, Sorgonà A, Ciaffi M. Diterpene Resin Acids and Olefins in Calabrian Pine (Pinus nigra subsp. laricio (Poiret) Maire) Oleoresin: GC-MS Profiling of Major Diterpenoids in Different Plant Organs, Molecular Identification and Expression Analysis of Diterpene Synthase Genes. Plants. 2021; 10(11):2391. https://doi.org/10.3390/plants10112391
Chicago/Turabian StyleAlicandri, Enrica, Stefano Covino, Bartolomeo Sebastiani, Anna Rita Paolacci, Maurizio Badiani, Francesco Manti, Carmelo Peter Bonsignore, Agostino Sorgonà, and Mario Ciaffi. 2021. "Diterpene Resin Acids and Olefins in Calabrian Pine (Pinus nigra subsp. laricio (Poiret) Maire) Oleoresin: GC-MS Profiling of Major Diterpenoids in Different Plant Organs, Molecular Identification and Expression Analysis of Diterpene Synthase Genes" Plants 10, no. 11: 2391. https://doi.org/10.3390/plants10112391