
Aspergillus niger Fermentation Broth Promotes Maize Germination and Alleviates Low Phosphorus Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Test Materials
2.1.2. Test Strain
2.1.3. Preparation of Fermentation Broth
2.1.4. Seed Germination Test
2.1.5. Seedling Hydroponic Experiment
2.2. Determination of Various Physiological and Germination Indices
2.2.1. Determination of Germination Rate and Germination Potential
2.2.2. Determination of Biomass
2.2.3. Determination of Root Morphological Index and Related Gene Expression
2.2.4. Determination of Chlorophyll Content
2.2.5. Determination of the Relative Water Content of Leaves and Soluble Sugar Content
2.2.6. Determination of Antioxidant Activity and Related Gene Expression
RNA Extraction and First Strand cDNA Synthesis
Real-Time Quantitative PCR
Primer Designing
2.3. Data Processing and Analysis
3. Results
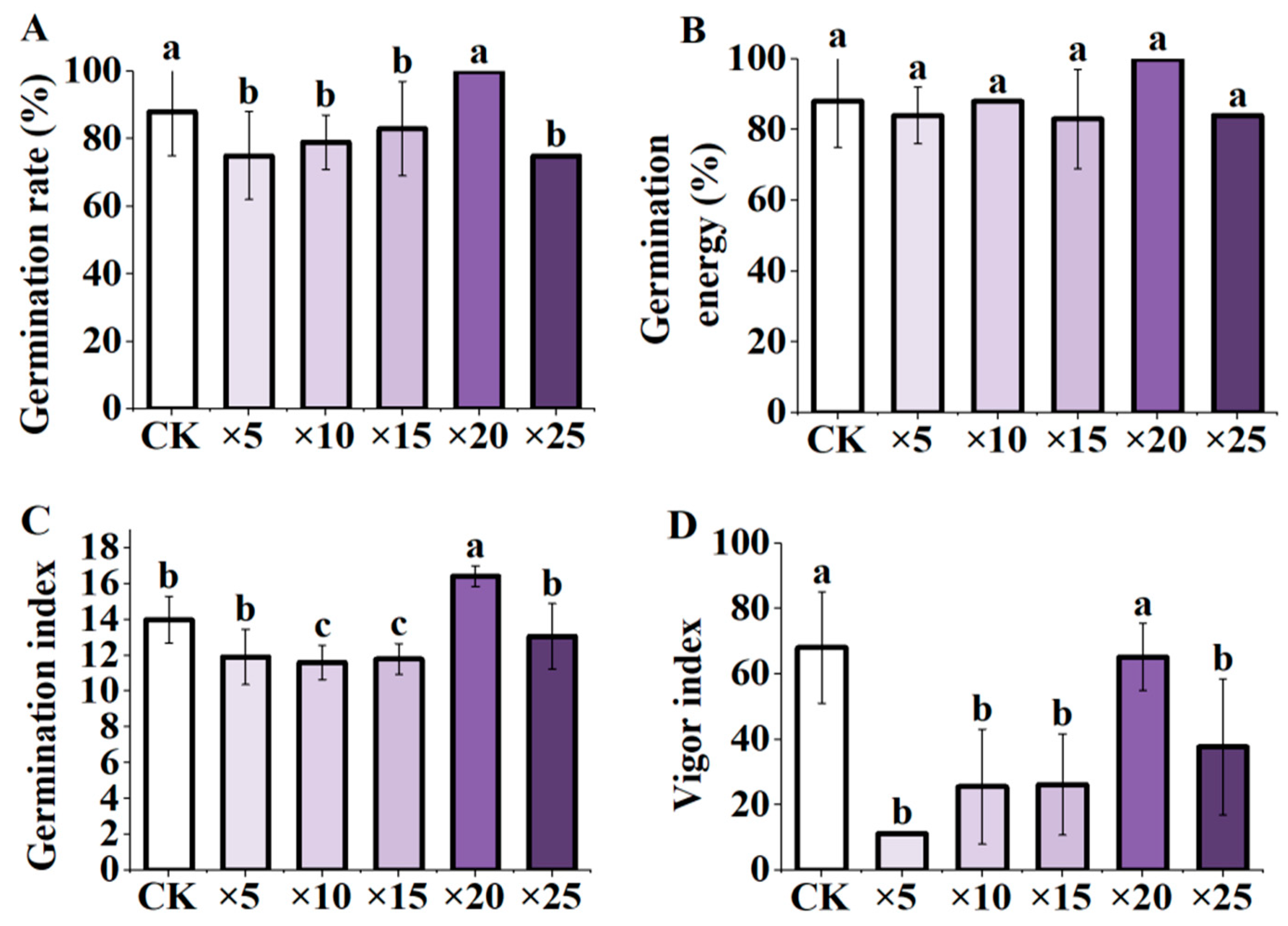
3.1. Effect of AFB Soaking on Maize Seed Germination Indexes under Different Dilution Ratios
3.2. The Effect of AFB on the Biomass of Maize Seedlings under Low Phosphorus Stress
3.3. Effects of AFB on Root Morphological Indicators under Low Phosphorus Stress
3.4. Effect of AFB on Chlorophyll Content of Maize Seedlings under Low Phosphorus Stress
3.5. Effects of AFB on Relative Water Content, MDA Content, and Soluble Sugar Content in Maize Seedling Leaves under Low Phosphorus Stress
3.6. Effects of AFB on Antioxidant Enzyme Activity in Leaves of Maize Seedlings under Low Phosphorus Stress
3.7. Effects of AFB on the Expression of Antioxidant Enzyme Activity-Related Genes in the Leaves of Maize Seedlings under Low Phosphorus Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AFB | Aspergillus niger fermentation broth |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| CAT | Catalase |
| MDA | Malondialdehyde |
| GI | Germination index |
| VI | Vigor index |
| GR | Germination rate |
| GE | Germination energy |
References
- Arif, M.; Shahzad, S.M.; Yasmeen, T.; Riaz, M.; Ashraf, M.; Ashraf, M.A.; Mubarik, M.S.; Kausar, R. Improving Plant Phosphorus (P) Acquisition by Phosphate-Solubilizing Bacteria. In Essential Plant Nutrients; Springer: Cham, Switzerland, 2017; pp. 513–556. [Google Scholar] [CrossRef]
- Pradhan, A.; Pahari, A.; Mohapatra, S.; Mishra, B.B. Phosphate-Solubilizing Microorganisms in Sustainable Agriculture: Genetic Mechanism and Application. Adv. Soil Microbiol. 2017, 2, 81–97. [Google Scholar] [CrossRef]
- Raghunandan, B.L. Rhizobacterial Phosphate Solubilizers in Sustainable Agriculture: Concepts and Prospects. Microorg. Green Revolut. 2017, 1, 107–124. [Google Scholar] [CrossRef]
- Kaur, G.; Reddy, M.S. Role of Phosphate-Solubilizing Fungi in Sustainable Agriculture. In Developments in Fungal Biology and Applied Mycology; Springer: Singapore, 2017; pp. 391–412. [Google Scholar] [CrossRef]
- Manoharachary, C.; Nagaraju, D.K. Role of Phosphate Solubilizing Fungi and Microbes for sustainable Agriculture and Agro forestry. Kavaka 2017, 48, 33–40. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi-current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Emami-Karvani, Z.; Chitsaz-Esfahani, Z. Phosphorus Solubilization: Mechanisms, Recent Advancement and Future Challenge. Sustain. Dev. Biodiversity. 2021, 27, 85–131. [Google Scholar] [CrossRef]
- Yuan, L.; Xihuan, L.; Ruixia, W.; Cai-ying, Z. Screen indexes for soybean tolerance to phosphorus deficiency and identification of low phosphorus tolerant soybean cultivars. J. Agric. Sci. Technol. 2015, 17, 30–41. [Google Scholar]
- Verlinden, M.S.; AbdElgawad, H.; Ven, A.; Verryckt, L.T.; Wieneke, S.; Janssens, I.A.; Vicca, S. Phosphorus stress strongly reduced plant physiological activity, but only temporarily, in a mesocosm experiment with Zea mays colonized by arbuscular mycorrhizal fungi. Biogeosciences 2022, 19, 2353–2364 . [Google Scholar] [CrossRef]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling Eutrophication: Nitrogen and Phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y.; et al. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef]
- Araújo, V.C.; Rossati, K.F.; Xavier, L.V.; Oliveira, V.A.; Carmo, G.J.; Assis, G.A.; Mendes, G.D. Enhanced growth in nursery of coffee seedlings inoculated with the rhizosphere fungus Aspergillus for field transplantation. Rhizosphere 2020, 15, 100236. [Google Scholar] [CrossRef]
- Jain, R.; Saxena, J.; Sharma, V. Effect of phosphate-solubilizing fungi Aspergillus awamori S29 on mungbean (Vigna radiata cv. RMG 492) growth. Folia Microbiol. 2012, 57, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Kuligod, V.; Hebsur, N.S.; Patil, C.R.; Kulkarni, G.N. Effect of phosphate solubilizing fungi and phosphorus levels on growth, yield and nutrient content in maize (Zea mays). Karnataka J. Agric. Sci. 2012, 25, 1. [Google Scholar]
- Kinjal, G.; TruptiPandya, J. A Review: Phosphate Solubilizing Microorganisms and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar]
- Pandya, N.; Desai, P.V.; Jadhav, H.P.; Sayyed, R.Z. Plant growth promoting potential of Aspergillus sp. NPF7, isolated from wheat rhizosphere in South Gujarat, India. Environ. Sustain. 2018, 1, 245–252. [Google Scholar] [CrossRef]
- Galeano, R.M.; Franco, D.G.; Chaves, P.O.; Giannesi, G.C.; Masui, D.C.; Ruller, R.; Corrêa, B.O.; da Silva Brasil, M.; Zanoelo, F.F. Plant growth promoting potential of endophytic Aspergillus niger 9-p isolated from native forage grass in Pantanal of Nhecolândia region, Brazil. Rhizosphere 2021, 18, 100332. [Google Scholar] [CrossRef]
- Wang, X.; Jarmusch, S.A.; Frisvad, J.C.; Larsen, T.O. Current status of secondary metabolite pathways linked to their related biosynthetic gene clusters in Aspergillus section Nigri. Nat. Prod. Rep. 2022, 40, 237–274. [Google Scholar] [CrossRef]
- Baker, S.E. Aspergillus niger genomics: Past, present and into the future. Med. Mycol. 2006, 44, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Bakri, M.M. Tri-Calcium and Zinc Phosphates Solubilization by Aspergillus niger and Its Relation to Organic Acids Production. Bionanoscience 2019, 9, 238–244. [Google Scholar] [CrossRef]
- Behera, B.C. Citric acid from Aspergillus niger: A comprehensive overview. Crit. Rev. Microbiol. 2020, 46, 727–749. [Google Scholar] [CrossRef]
- Mendoza-Arroyo, G.E.; Chan-Bacab, M.J.; Aguila-Ramírez, R.N.; Ortega-Morales, B.O.; Canché Solís, R.E.; Chab-Ruiz, A.O.; Cob-Rivera, K.I.; Dzib-Castillo, B.B.; Tun-Che, R.E.; Camacho-Chab, J.C. Inorganic Phosphate Solubilization by a Novel Isolated Bacterial Strain Enterobacter sp. ITCB-09 and Its Application Potential as Biofertilizer. Agriculture 2020, 10, 383. [Google Scholar] [CrossRef]
- Sugiharto, A.; Napitupulu, T.P.; Sudiana, M.I. The influence of biocarrier of Aspergillus niger and Trichoderma harzianum toward vegetative growth of sorghum in the field experiment. J. Microb. Syst. Biotechnol. 2020, 2, 28–34. [Google Scholar] [CrossRef]
- Medina, A.; Vassileva, M.; Barea, J.M.; Azcón, R. The growth-enhancement of clover by Aspergillus-treated sugar beet waste and Glomus mosseae inoculation in Zn contaminated soil. Appl. Soil Ecol. 2006, 33, 87–98. [Google Scholar] [CrossRef]
- Fan, J.; Wei, G.; Zhang, H.; Zhao, W.; Qiu, H.; Hu, A.; Li, Z. Effects of self-developed Aspergillus niger pelletized seed coating on sorghum seed germination and seedling growth. Acta Agric. Univ. Jiangxiensis 2021, 1, 25–32. [Google Scholar] [CrossRef]
- Naeem, U.; Haq, I.U.; Afzaal, M.; Qazi, A.; Yasar, A.; Tabinda, A.B.; Mahfooz, Y.; Naz, A.U.; Awan, H.A. Investigating the effect of Aspergillus niger inoculated press mud (biofertilizer) on the potential of enhancing maize (Zea mays. L.) yield, potassium use efficiency and potassium agronomic efficiency. Cereal Res. Commun. 2021, 50, 157–170. [Google Scholar] [CrossRef]
- Vassilev, N.; Vassileva, M.; Lopez, A.; Martos, V.; Reyes, A.; Maksimovic, I.; Eichler-Löbermann, B.; Malusà, E. Unexploited potential of some biotechnological techniques for biofertilizer production and formulation. Appl. Microbiol. Biotechnol. 2015, 99, 4983–4996. [Google Scholar] [CrossRef]
- Zhang, H. Analysis and Prospect of China’s Corn Production Cost Status. J. Soc. Sci. Humanit. 2022, 4, 1–3. [Google Scholar] [CrossRef]
- Marković, M.; Komljenović, I.; Kovačević, V.; Radić, V.; Jović, J.; Trbic, G.; Pesevic, D.; Marković, M. Alleviation of Negative Climate Change Effects on Maize Yields in Northern Bosnia by Liming and Phosphorus Fertilization. In Climate Change Adaptation in Eastern Europe; Springer: Cham, Switzerland, 2019; pp. 169–183. [Google Scholar] [CrossRef]
- Li, D.; Wang, H.; Wang, M.; Li, G.; Chen, Z.; Leiser, W.L.; Weiß, T.M.; Lu, X.; Wang, M.; Chen, S.; et al. Genetic Dissection of Phosphorus Use Efficiency in a Maize Association Population under Two P Levels in the Field. Int. J. Mol. Sci. 2021, 22, 9311. [Google Scholar] [CrossRef]
- Yuan, H.; Chen, H.; Gao, D.; Liu, X.; Guo, B.; Li, Z. Determination of Aspergillus niger spores concentration by spectrophotometry. China Brewing 2017, 4, 122–126. [Google Scholar] [CrossRef]
- Li, Z.; Ge, Y.; Chen, Q.; Zhou, L.; Liu, W. Antifungal Activity and Stability of Aspergillus niger xj Fermentation Broth. Food Res. Dev. 2011, 7, 141–143. [Google Scholar]
- Huang, Z.; Zhi, J.; Li, L.; Zheng, Y.; He, L. Study on the Revision of the Botanical Name of the “Rules for Agricultural Seed Testing”. Seed 2020, 3, 105–119. [Google Scholar] [CrossRef]
- Morales-Quintana, L.; Moya, M.; Santelices-Moya, R.E.; Cabrera-Ariza, A.; Rabert, C.; Pollmann, S.; Ramos, P. Improvement in the physiological and biochemical performance of strawberries under drought stress through symbiosis with Antarctic fungal endophytes. Front. Microbiol. 2022, 13, 939955. [Google Scholar] [CrossRef]
- Mulati, N.; Yang, Y.; Abdukrimu, P.; Wusman, M. Comparison of Determination Methods for Chlorophyll Content in Wheat. Jiangsu Agric. Sci. 2021, 9, 156–159. [Google Scholar] [CrossRef]
- Liang, Y.; Shi, Y.; Jin, X.; Zhang, S.; Hou, L.; Zhang, Y. Effects of Light Quality on the Growth and Physiological Resistance of Tomato Seedlings Under Low- Phosphorus Stress. Chin. Agric. Sci. Bull. 2020, 1, 56–61. [Google Scholar]
- Weatherley, P.E. Studies in the water relations of the cotton plant 1. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Zhang, S.; Zong, Y.; Fang, C.; Huang, S.; Li, J.; Xu, J.; Wang, Y.; Liu, C. Optimization of Anthrone Colorimetric Method for Rapid Determination of Soluble Sugar in Barley Leaves. Food Res. Dev. 2020, 7, 196–200. [Google Scholar] [CrossRef]
- Draper, H.H.; Squires, E.J.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free. Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Zheng, S.; Shang, X.; Wang, J. Determination of anti-oxidant enzyme activity and malondialdehyde content in maize seedlings under salt stress by visible spectrophotometry. Biotechnol. Bull. 2010, 7, 106–109. [Google Scholar] [CrossRef]
- Rehmani, M.S.; Aziz, U.; Xian, B.; Shu, K. Seed Dormancy and Longevity: A Mutual Dependence or a Trade-Off? Plant Cell Physiol. 2022, 63, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Rehmani, M.S.; Xian, B.; Wei, S.; He, J.; Feng, Z.; Huang, H.; Shu, K. Seedling establishment: The neglected trait in the seed longevity field. Plant Physiol. Biochem. PPB 2023, 200, 107765. [Google Scholar] [CrossRef]
- Basuchaudhuri, P. Seed Germination. In Physiology of Soybean Plant, 1st ed.; CRC Press: Boca Raton, FL, USA, 2020; 32p. [Google Scholar] [CrossRef]
- Lv, J.; Tian, X.; Yan, H.; Cao, C. Effect of Aspergillus niger fermentation liquid on the growth of wheat seedlings in calcareous soil. J. Northwest AF Univ. 2015, 5, 100–106. [Google Scholar] [CrossRef]
- Yue, Z.; Shen, Y.; Chen, Y.; Liang, A.; Chu, C.; Chen, C.; Sun, Z. Microbiolog-ical Insights into the Stress-Alleviating Property of an Endophytic Bacillus altitudinis WR-10 in Wheat under Low-Phosphorus and High-Salinity Stresses. Microorganisms 2019, 7, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Yang, J.; Chen, Q.; Zhang, Y.; Zhang, Y.; Feng, J.; Jia, M. Research on Effects of AMF Addition and Plants Combinations on Growth and Uranium Enrichment of Lolium perenne. Nonferrous Met. 2022, 1, 108–117. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, H.; Liu, W.; Li, Z.; Chen, Q. Analysis of the fatty acid from Aspergillus Niger XJ fermentation broth. Food Eng. 2012, 123, 31–33. [Google Scholar]
- Li, C.; Tian, X.; Cao, C. Effects of Exogenous Organic Acids on Phosphate Uptake and Growth of Maize. Acta Bot. Boreali Occident. Sin. 2011, 31, 1376–1383. [Google Scholar]
- Wu, Q.; Hu, H.; He, L. Effect of Aspergillus niger Strain Xf-1 on Soil Nutrients and Growth of Amorpha Fruticosa. Appl. Ecol. Environ. Res. 2020, 18, 5079–5092. [Google Scholar] [CrossRef]
- Nuangmek, W.; Aiduang, W.; Kumla, J.; Lumyong, S.; Suwannarach, N. Evaluation of a Newly Identified Endophytic Fungus, Trichoderma phayaoense for Plant Growth Promotion and Biological Control of Gummy Stem Blight and Wilt of Muskmelon. Front. Microbiol. 2021, 12, 634772. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, W.; Yan, Y.; Si-Yu, G. Using sturdy-seeding agent which inoculates fungi of dissolving phosphorus to the influence of rice seedling growth characteristic and phosphorus absorption. Soil Fertil. Sci. China 2008, 213, 69–72. [Google Scholar]
- Rahmansyah, M.; Sugiharto, A.; Juhaeti, T. Effect of Aspergillus niger inoculant to sorghum seedling growth due to drought stress and its growth in the field. Pros. Semin. Nas. Masy. Biodiversitas Indones. 2017, 3, 426–432. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, L.; Huang, L.; Sun, X.; Wang, P. Response of growth characteristics and endogenous hormones of Sophora davidii to low-phosphorus stress. Acta Physiol. Plant. 2021, 43, 1–14. [Google Scholar] [CrossRef]
- Li, P.; Weng, J.; Rehman, A.; Niu, Q. Root Morphological and Physiological Adaptations to Low Phosphate Enhance Phosphorus Efficiency at Melon (Cucumis melo L.) Seedling Stage. Horticulturae 2022, 8, 636. [Google Scholar] [CrossRef]
- Yahya, M.; Islam, E.U.; Rasul, M.; Farooq, I.; Mahreen, N.; Tawab, A.; Irfan, M.; Rajput, L.B.; Amin, I.; Yasmin, S. Differential Root Exudation and Architecture for Improved Growth of Wheat Mediated by Phosphate Solubilizing Bacteria. Front. Microbiol. 2021, 12, 744094. [Google Scholar] [CrossRef]
- Mundim, G.S.; Maciel, G.M.; Mendes, G.D. Aspergillus niger as a Biological Input for Improving Vegetable Seedling Production. Microorganisms 2022, 10, 674. [Google Scholar] [CrossRef]
- Lubna Asaf, S.; Hamayun, M.; Gul, H.; Lee, I.; Hussain, A. Aspergillus niger CSR3 regulates plant endogenous hormones and secondary metabolites by producing gibberellins and indoleacetic acid. J. Plant Interact. 2018, 13, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, A.; Hussain, A.; Irshad, M.; Hamayun, M.; Iqbal, A.; Khan, N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis 2018, 77, 225–235. [Google Scholar] [CrossRef]
- Chen, B.; Wang, Q.; Bücking, H.; Sheng, J.; Luo, J.; Chai, Z.; Kafle, A.; Hou, Y.Y.; Feng, G. Genotypic differences in phosphorus acquisition efficiency and root performance of cotton (Gossypium hirsutum) under low-phosphorus stress. Crop Pasture Sci. 2019, 70, 344–358. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, anti-oxidants and stress tolerance. Trends Plant Sci. 2002, 79, 405–410. [Google Scholar] [CrossRef]
- Khuna, S.; Suwannarach, N.; Kumla, J.; Frisvad, J.C.; Matsui, K.; Nuangmek, W.; Lumyong, S. Growth Enhancement of Arabidopsis (Arabidopsis thaliana) and Onion (Allium cepa) With Inoculation of Three Newly Identified Mineral-Solubilizing Fungi in the Genus Aspergillus Section Nigri. Front. Microbiol. 2021, 12, 705896. [Google Scholar] [CrossRef]
- Abdel-Motaal, F.F.; Kamel, N.M.; El-zayat, S.A.; Abou-Ellail, M. Early blight suppression and plant growth promotion potential of the endophyte Aspergillus flavus in tomato plant. Ann. Agric. Sci. 2020, 65, 117–123. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Ghaderi, N.; Havas, F.; Dedejani, S. Comparative investigation of structural relationships among morpho-physiological and biochemical properties of strawberry (Fragaria × ananassa Duch.) under drought and salinity stresses: A study based on in vitro culture. Sci. Hortic. 2019, 256, 108601. [Google Scholar] [CrossRef]
- Zhang, Y.; Luan, Q.; Jiang, J.; Li, Y. Prediction and Utilization of Malondialdehyde in Exotic Pine Under Drought Stress Using Near-Infrared Spectroscopy. Front. Plant Sci. 2021, 12, 735275. [Google Scholar] [CrossRef]
- Han, Y.; Fan, S.; Zhang, Q.; Wang, Y. Effect of heat stress on the MDA, proline and soluble sugar content in leaf lettuce seedlings. Agric. Sci. 2013, 4, 112–115. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimian, E.; Bybordi, A. Influence of ascorbic acid foliar application on chlorophyll, flavonoids, anthocyanin and soluble sugar contents of sunflower under conditions of water deficit stress. J. Food Agric. Environ. 2012, 10, 1026–1030. [Google Scholar]
- Ding, Z.; Hu, N.; Zhu, R.; Ma, J.; Sun, J.; Li, G. Synergistic Enhancement of from Soil by Aspergillus niger fermentation broth and Calcium peroxide. Nonferrous Met. Eng. 2020, 11, 129–136. [Google Scholar] [CrossRef]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Singh, H.P.; Cha-Um, S. Foliar application of glycinebetaine regulates soluble sugars and modulates physiological adaptations in sweet potato (Ipomoea batatas) under water deficit. Protoplasma 2020, 257, 197–211. [Google Scholar] [CrossRef]
- Gao, Y.; Long, R.; Kang, J.; Wang, Z.; Zhang, T.; Sun, H.; Li, X.; Yang, Q. Comparative Proteomic Analysis Reveals That Anti-oxidant System and Soluble Sugar Metabolism Contribute to Salt Tolerance in Alfalfa (Medicago sativa L.) Leaves. J. Proteome Res. 2019, 18, 191–203. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Chen, J.; Li, Z.; Liu, W.; Jiang, X.; Yu, S. Effects of Arbuscular Mycorrhizal Fungi on the Growth and Physiology of Cinnamomum camphora(L.) Seedlings Under Pb Stress. Acta Agric. Univ. Jiangxiensis 2021, 3, 621–629. [Google Scholar] [CrossRef]
- Ahmad, P.; Ashraf, M.; Hakeem, K.R.; Azooz, M.M.; Rasool, S.; Chandna, R.; Akram, N.A.; Arabia, S. Potassium starvation-induced oxidative stress and anti-oxidant defense responses in Brassica juncea. J. Plant Interact. 2012, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Erdoğan, Ü.; Çakmakçı, R.; Varmazyari, A.; Turan, M.; Erdoğan, Y.; Kitir, N. Role of inoculation with multi-trait rhizobacteria on strawberries under water deficit stress. Zemdirbyste-Agriculture 2016, 103, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front. Plant Sci. 2016, 7, 01787. [Google Scholar] [CrossRef] [Green Version]
- Sekmen Cetinel, A.H.; Gokce, A.; Erdik, E.; Çetinel, B.; Cetinkaya, N. The Effect of Trichoderma citrinoviride Treatment under Salinity Combined to Rhizoctonia solani Infection in Strawberry (Fragaria × ananassa Duch.). Agronomy 2021, 11, 1589. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M. Potassium upregulates anti-oxidant metabolism and alleviates growth inhibition under water and osmotic stress in wheat (Triticum aestivum L.). Protoplasma 2016, 254, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, Z.; Ghanizadeh, H.; Kerckhoffs, H.; Sofkova-Bobcheva, S.; Wu, W.; Wang, X.; Liu, Y.; Li, X.; Zhao, H.; et al. Comparative Genomic and Physiological Analyses of a Superoxide Dismutase Mimetic (SODm-123) for Its Ability to Respond to Oxidative Stress in Tomato Plants. J. Agric. Food Chem. 2020, 68, 13608–13619. [Google Scholar] [CrossRef] [PubMed]
- Pawar, V.C.; Thaker, V.S. Acid phosphatase and invertase activities of Aspergillus niger. Mycoscience 2009, 50, 323–330. [Google Scholar] [CrossRef]
- Begum, N.; Wang, L.; Ahmad, H.; Akhtar, K.; Roy, R.; Khan, M.I.; Zhao, T. Co-inoculation of Arbuscular Mycorrhizal Fungi and the Plant Growth-Promoting Rhizobacteria Improve Growth and Photosynthesis in Tobacco Under Drought Stress by Upregulating Antioxidant and Mineral Nutrition Metabolism. Microb. Ecol. 2021, 83, 971–988 . [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primers (5′-3′) | Reverse Primers (3′-5′) | |
|---|---|---|
| SOD9 | GGCTGTTGCTGTGCTTGGTA | CTTGCTCGCAGGATTGTAGTG |
| CAT2 | CCCCAACTACCTGCTGCTAC | TGGTTATGAACCGCTCTTGC |
| LOX3 | CGGCGTTCAAGAGGTTCAG | TGGTCAGAGGTGTTGGGATAGA |
| EXPB4 | CAAGCACACCAACCAGTA | GCACCGAATCTTGTAGCA |
| KRP1 | CCGTATCTCCAGCCATTG | GCCGAGGACCTAGAACAT |
| GADPH | CCATCACTGCCACACAGAAAAC | AGGAACACGGAAGGACATACCAG |
| Treatment | Total Length (cm) | SurfArea (cm2) | ProjArea (cm2) | AvgDiam (mm) | RootVolume (cm3) | Tips | Forks |
|---|---|---|---|---|---|---|---|
| CK | 658.76 ± 151.66 bc | 318.53 ± 47.69 b | 101.39 ± 15.18 b | 1.57 ± 0.18 c | 12.38 ± 1.61 b | 2874 ± 513 c | 6081 ± 2412 b |
| CK+A | 589.14 ± 89.95 c | 353.33 ± 67.91 b | 112.47 ± 21.62 b | 1.87 ± 0.14 c | 15.09 ± 1.91 b | 1681 ± 402 d | 3261 ± 175 c |
| P | 884.05 ± 184.8 b | 415.61 ± 43.51 b | 132.29 ± 13.85 b | 2.93 ± 0.23 b | 15.67 ± 0.81 b | 6580 ± 556 b | 8723 ± 828 b |
| P+A | 1165.63 ± 171.17 a | 546.50 ± 92.94 a | 173.96 ± 29.58 a | 3.64 ± 0.76 a | 22.3 ± 3.75 a | 9602 ± 1247 a | 12,208 ± 1596 a |
| Treatment | Total Chlorophyll | Chlorophyll a | Chlorophyll b | Carotenoids |
|---|---|---|---|---|
| CK | 3.33 ± 0.28 c | 2.17 ± 0.26 c | 1.51 ± 0.09 b | 0.17 ± 0.03 c |
| CK+A | 2.72 ± 0.33 c | 2.01 ± 0.2 c | 1.43 ± 0.08 b | 0.18 ± 0.01 c |
| P | 7.82 ± 0.61 b | 5.93 ± 0.54 b | 3.52 ± 0.49 a | 0.45 ± 0.05 b |
| P+A | 8.99 ± 0.57 a | 6.82 ± 0.45 a | 3.5 ± 0.21 a | 0.66 ± 0.07 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, M.; Zhang, C.; Zhang, Z.; Jiang, T.; Hu, X.; Qiu, H.; Li, Z. Aspergillus niger Fermentation Broth Promotes Maize Germination and Alleviates Low Phosphorus Stress. Microorganisms 2023, 11, 1737. https://doi.org/10.3390/microorganisms11071737
Tian M, Zhang C, Zhang Z, Jiang T, Hu X, Qiu H, Li Z. Aspergillus niger Fermentation Broth Promotes Maize Germination and Alleviates Low Phosphorus Stress. Microorganisms. 2023; 11(7):1737. https://doi.org/10.3390/microorganisms11071737
Chicago/Turabian StyleTian, Maoxian, Changhui Zhang, Zhi Zhang, Tao Jiang, Xiaolan Hu, Hongbo Qiu, and Zhu Li. 2023. "Aspergillus niger Fermentation Broth Promotes Maize Germination and Alleviates Low Phosphorus Stress" Microorganisms 11, no. 7: 1737. https://doi.org/10.3390/microorganisms11071737