
Five Species of Taxus Karyotype Based on Oligo-FISH for 5S rDNA and (AG3T3)3

Southwest Engineering Technology Research Center of Taxus of State Forestry and Grassland Administration, College of Forestry, Sichuan Agricultural University, Huimin Road 211, Chengdu 611130, China
*
Author to whom correspondence should be addressed.
Genes 2022, 13(12), 2209; https://doi.org/10.3390/genes13122209
Submission received: 27 September 2022
/
Revised: 7 November 2022
/
Accepted: 21 November 2022
/
Published: 25 November 2022
(This article belongs to the Special Issue Chromosome Evolution and Karyotype Analysis)
Abstract
:As a relict plant, Taxus is used in a variety of medicinal ingredients, for instance to treat a variety of cancers. Taxus plants are difficult to distinguish from one another due to their similar morphology; indeed, some species of Taxus cytogenetic data still are unclear. Oligo-FISH can rapidly and efficiently provide insight into the genetic composition and karyotype. This is important for understanding the organization and evolution of chromosomes in Taxus species. We analysed five Taxus species using two oligonucleotide probes. (AG3T3)3 signals were distributed at the chromosome ends and the centromere of five species of Taxus. The 5S rDNA signal was displayed on two chromosomes of five species of Taxus. In addition to Taxus wallichiana var. mairei, 5S rDNA signals were found proximal in the remaining four species, which signals a difference in its location. The karyotype formula of Taxus wallichiana was 2n = 2x = 24m, its karyotype asymmetry index was 55.56%, and its arm ratio was 3.0087. Taxus × media’s karyotype formula was 2n = 2x = 24m, its karyotype asymmetry index was 55.09%, and its arm ratio was 3.4198. The karyotype formula of Taxus yunnanensis was 2n = 2x = 24m, its karyotype asymmetry index was 55.56%, and its arm ratio was 2.6402. The karyotype formula of Taxus cuspidate was 2n = 2x = 24m, its karyotype asymmetry index was 54.67%, its arm ratio was 3.0135, and two chromosomes exhibited the 5S rDNA signal. The karyotype formula of T. wallichiana var. mairei was 2n= 2x = 22m + 2sm, its karyotype asymmetry index was 54.33%, and its arm ratio was 2.8716. Our results provide the karyotype analysis and physical genetic map of five species of Taxus, which contributes to providing molecular cytogenetics data for Taxus.
1. Introduction
Taxus L. is an ancient tertiary relict plant [1] rich in substances such as biflavones, cinnamic acid derivatives, lignans, and steroids [2,3]. In particular, it is rich in the natural taxane diterpene paclitaxel, which, along with some semisynthetic drugs, is used to treat ovarian, breast, skin, and non-small cell lung cancer [4,5]. For the protection of wild Taxus plants, the Convention on International Trade in Endangered Species (CITES) has listed all Taxus species in Appendix II. Subsequently, all Taxus species in China were included in the National List of Key Protected Wild Plants (http://www.Gov.cn/Baogong/content/2000/content_60072.htm (accessed on 2 September 2022)). Therefore, it is of great significance to study Taxus.
Taxus is an important part of Taxaceae, distributed in the northern hemisphere such as in Europe, North America, and East Asia, and it is suitable for growing under humid temperate or tropical mountain forests [1,6,7,8,9]. There are four species and one variety in China, distributed in the northeast, southwest, and south regions [10]. Taxus classification is mainly based on morphological description and geographical distribution [11], but some species of Taxus have little difference in phenotypic characteristics [9], causing the controversial classification of species of Taxus. Its phylogeny remains controversial [7]; some researchers have divided Taxus into 7 [12], 8 [13], 9 [14], 10 [10], 10 species and their varieties [11] and 11 [15] species. Taxus is classified only by appearance, which is limited and therefore makes it easy to cause classification difficulties. We are looking forward to a more reasonable method. At present, the nuclear and chloroplast genomes of Taxus wallichiana var. mairei have been sequenced [16] and the chloroplast (cp) genome of Taxus chinensis has been sequenced [16]. The mitochondrial (mt) genome of Taxus cuspidate has also been sequenced [17]. Taxus as a non-model gymnosperm has been studied for organ transcriptome sequencing at present [18]. A draft genome assembly constructed by the whole genome sequencing of Taxus baccata has been published [19]. Although some molecular studies have been conducted on certain Taxus species [20,21], the whole genome sequencing of Taxus is still missing [22]. One of the ways in which plant genomic diversity is expressed is through a wide range of chromosome numbers [23]. However, there are still significant gaps in the overall study of the major cytogenetic characteristics of the Taxaceae genome [22]. Compared with other traits, chromosomal traits are stably informative in plants [24]. Karyotypes mainly contain chromosome numbers and morphology, which reveal cytological characteristics [25]. It is better to understand the organization and evolution of chromosomes in related species [26].
Fluorescence in situ hybridization (FISH) is an important tool for plant karyotypes. which uses oligonucleotide probes [27]. FISH also frequently uses oligonucleotide DNA and other DNA sequences as probes, and has been used on a variety of plants, such as Pinus densiflora [28], Chimonanthus campanulatus [29], Citrus clementina [30,31,32], and Pinus [33,34,35]. Telomere repeats (CCCTAAA)n, (TTTAGGG)n, (TTAGGG)n, and (AGGGTTT)n are found in most angiosperms and are located at the end of chromosomes in gymnosperms. Picea asperata and Larix gmelinii show only terminal sites, but in Picea sylvestris [36,37] and Picea elliottii, several insertion sites at the centromere sites were also observed [38]. The 5S rDNA and (AG3T3)3 probe combination has been applied to Hibiscus mutabilis [39], Hippophae rhamnoides [40], and C. campanulatus [29]. There is no systematic karyotype analysis of Taxus, and the results of previous karyotype studies often have some differences because of the different methods used. To study the karyotypes of five Taxus species, we used two oligonucleotide probes that have not yet been used. Our results help to distinguish five Taxus species based on chromosome information, and these results will be helpful for the construction of oligonucleotide barcodes in the future.
2. Materials and Methods
2.1. Seed Materials and Root Tips
The seedlings of Taxus yunnanensis, Taxus × media, and T. wallichiana var. mairei. were collected in Caoba town, Yucheng district, Ya’an City, Sichuan province. Taxus wallichiana was collected from Xichang city, Sichuan province. Taxus cuspidate was collected in Suqian city, Jiangsu province. All five cultivated Taxus species are shown in Table 1. For each Taxus species, there are three healthy seedlings, each with seeds from the same plant for consistent genotype. The Taxus seedlings had been growing for two years before the experiment. The seedling morphology and root tip growth and division of each Taxus species were similar. These seedlings were placed at room temperature under natural light conditions in the Chengdu Academy of Agriculture and Forestry Sciences. Approximately 1.5–2.0 cm root tips were treated with nitrous oxide for 4.5 h. After treatment, the root tips were placed in glacial acetic acid for 5 min and then maintained in 75% ethyl alcohol at −20 °C.
2.2. Chromosome Preparation
The root tip meristems were washed by using ddH2O, and then dispersed with cellulose and pectinase (2:1) at 37 °C for 45 min. After these treatments, the enzyme mixture on the meristems was washed off using ddH2O twice, then ethyl alcohol twice. Then, we removed all ethyl alcohol and added up to 20 µL of glacial acetic acid to prepare a suspension. Finally, 10 µL of the mixture was dropped on one slide. We used an Olympus CX23 microscope (Olympus Corporation, Tokyo, Japan) to examine the slides.
2.3. Probe Preparation
The (AG3T3)3 repeat sequence [24] and the ribosome 5S rDNA [41] were used in this study. These two probes were tested for the first time in Taxaceae and these oligonucleotide sequences were produced by Sangon Biotechnology Co., Ltd. (Shanghai, China). The 5′ ends of the probes were labelled with 6-carboxyfluorescein (FAM) or 6-carboxytetramethylrhodamine (TAMRA). The probes were dissolved in 1 × Tris-ethylene diamine tetra acetic acid (TE) and maintained at a concentration of 10 µM at −20 °C.
2.4. FISH Hybridization
The slides were fixed in 4% paraformaldehyde for 10 min and oscillated twice in 2 × saline sodium citrate (SSC) buffer for 5 min. After, the samples were dipped in 75%, 95%, and 100% ethyl alcohol successively for 5 min. A total of 60 µL of 70% deionized formamide (FA) was dropped onto the slides and air dried, and coverslips (24 cm × 50 cm) were placed in 70% FA at 80 °C for 2 min. We removed the coverslips from the slides, and the slides were then incubated in 75%, 95%, and 100% ethanol (precooled at 20 °C) for 5 min. A total of 10 µL of hybridization solution, including 0.5 µL of the (AG3T3)3 probe, 1 µL of 5S rDNA, and 8.5 µL of the 2 × SSC, and 1 × TE mixture was dropped onto the chromosomes, and a cover glass (24 cm × 50 cm) was placed on top of the hybrid solution. The slides were then incubated at 37 °C for 2 h.
2.5. Image Capture and Analysis
The slides with hybridization were successively washed with 2 × SSC buffer and ddH2O. A total of 10 µL of 4,6-diamidino-2-phenylindole (DAPI) was dropped onto the air-dried chromosomes, then coverslips (24 cm × 50 cm) were placed on top. The slides were examined using an Olympus BX 63 fluorescence microscope combined with a Photometric SenSys Olympus DP70 CCD camera (Olympus Corporation, Tokyo, Japan).
We conducted an analysis of the signal patterns by using the three best spreads. We used Photoshop version 2021 (Adobe Systems Inc., San Jose, CA, USA) to calculate the length of each chromosome, and each spread was measured three times, which obtained consistent chromosome data. The chromosomes were arranged by length from longest to shortest.
3. Results
3.1. Morphological Characteristics
T. wallichiana leaves were striped, more densely arranged into two irregular columns overlapping each other, thick in texture, and usually straight. Some were equal in width at the top or bottom or slightly narrow at the upper end and were symmetrical at the base.
The leaves of T. × media were arranged in two irregular rows or slightly spirally. The leaves were striped, falcate curved, dense green, and slightly raised in the middle rib.
T. yunnanensis’ leaf texture was thin and soft, and the shape of the leaf was striped lanceolate.
T. cuspidate leaves were arranged in two irregular rows that were observably extending at an angle of about 45 degrees. Their leaves were bar-shaped, usually straight, thinly and slightly curved, narrow at the base, and had a short stripe.
T. wallichiana var. mairei leaves were often wider and longer, the upper part was often narrow, the apex was acuminate, and the midvein belt was clearly visible. The leaves were yellowish green or green, and the green sideband was also wider and obvious.
The leaf and bud morphology of these five plants are shown in Figure 1.
3.2. Chromosome
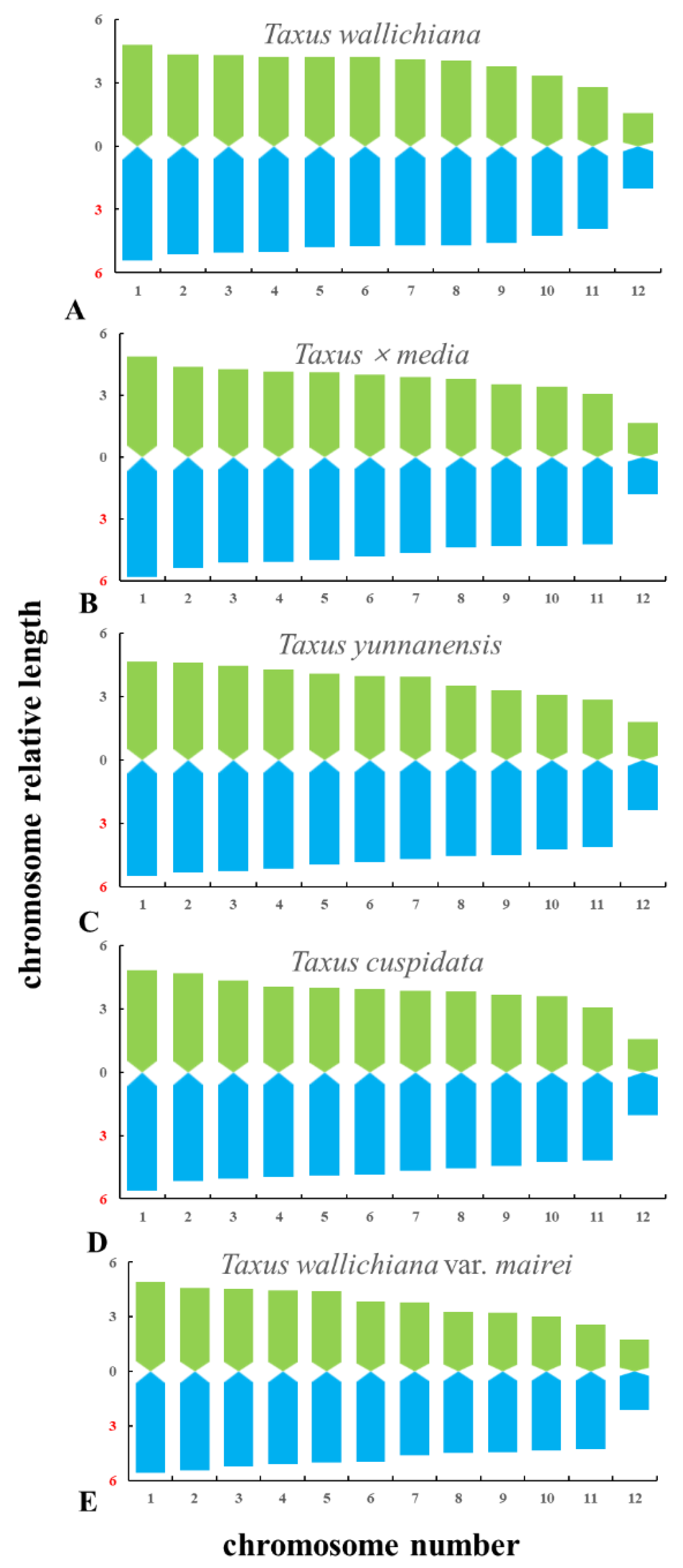
The metaphase and prometaphase chromosomes of five species of Taxus were analysed by FISH, as shown in Figure 2. The chromosome number of all five species of Taxus was 2n = 24. The individual chromosomes in Figure 2 were aligned based on length from the longest chromosome to the shortest chromosome, as shown in Figure 3. Except for the different karyotype formula of T. wallichiana var. mairei, the other four Taxus species have the same karyotype formula. The arm ratios of five Taxus species range from 2.6402 to 3.4198. The karyotype asymmetry index ranges from 54.33% to 55.56%. The karyotypes of the five Taxus species are shown in Table 2. The relative chromosome lengths of the five species of Taxus are shown in Figure 4.
3.3. Signal Distribution
The (AG3T3)3 signal on chromosomes of five species of Taxus appeared at the centromere and proximal. The (AG3T3)3 signal showed slight differences, but it may help to count the chromosome number of the five species of Taxus. T. cuspidate and T. × media had two pairs of 5S rDNA signals, and the (AG3T3)3 signal location was shown to be in the centromere and chromosomal endpoint. The 5S rDNA signals of T. wallichiana and T. yunnanensis were distributed in centromeres and subtelomeres, and the number of 5S rDNA signals was consistent. The (AG3T3)3 signal of T. wallichiana and T. yunnanensis was distributed at the chromosome end. The 5S rDNA presented two signals at different locations in five species of Taxus, showing a great ability for chromosome discrimination. The 5S rDNA signals of T. wallichiana and T. yunnanensis were similar, but the individual 5S rDNA signal location of T. yunnanensis was more in the centre than in T. wallichiana. The 5S rDNA signals of T. × media and T. cuspidate were similar, but the (AG3T3)3 signals of T. × media in chromosomes 17 and 18 were fewer than in T. cuspidate, as shown in Figure 5.
4. Discussion
4.1. Karyotype
Taxus, as a non-model gymnosperm, has been studied for organ transcriptome sequencing [18,42,43]. A draft genome assembly constructed by the whole genome sequencing of T. baccata has been published [19]. Although some molecular studies have been conducted on certain Taxus species [20,21], the whole genome sequencing of Taxus is still missing [22]. Although the complete genome sequencing of Taxus is missing, plant genomic diversity can also be reflected by a wide range of chromosome numbers [43]. Therefore, studying the chromosomes of five species of Taxus can also provide reference for genome research, to a certain extent.
Compared with angiosperms, gymnosperms are characterized by more consistent chromosome numbers and karyotypes [43]. Gymnosperms are one of the best studied plants in terms of chromosome number and karyotype [22,23,44,45]. To date, the Taxus family is the only gymnospermic family known to produce paclitaxel [46]. However, information on the Taxus family itself is limited, because the identification of the Taxus chromosomes is complicated due to their similar size and morphology [22].
In this study, chromosome numbers were the same as those previously published [47,48]. However, the karyotype formulas of the five species of Taxus were different from those previously published. In this study, except for the karyotype formula of T. wallichiana var. mairei being 2n = 2X = 22m + 2sm, the karyotype formulas of the other four Taxus are 2n = 2X = 24m. The previous study showed that the karyotype formulas of male T. yunnanensis were 2n = 24 = 24m, and those for females were 2n = 24 = 23m + 1sm [47] and 2n = 24 = 21m + 1sm + 2T [47]. The karyotype formula of T. × media was 2n = 24 = 20m + 2sm + 2st [48]. The karyotype formulas of T. cuspidata were 2n = 24 = 18m + 6sm [49], 2n = 24 = 18m + 4sm + 2T [48], and 2n = 24 = 4m + 6msm + 10m + 2sm + 2T [50]. The karyotype formula of T. wallichiana var. mairei was 2n = 24 = 20m + 2sm + 2T [47].
The karyotype formulas of different species are varied, which may be due to cell cycle synchronization and low chromosomal diffusion efficiency [22] or could be due to the differential accumulation of transposable factors [51,52]. The different treatment methods of Taxus root tip materials may also result in the inconsistency between the karyotype formula and previous studies.
4.2. Role of (AG3T3)3 and 5S rDNA
By using the FISH technique, the (AG3T3)3 probe was used on five species of Taxus, and signals were found on all chromosomes, indicating that the chromosome shape of five species of Taxus was complete. Traditionally, (AG3T3)3 signals are usually detected at the chromosome end, but occasionally some signals are detected around and inside the centromere [31,53,54,55]. Studies have shown that the presence of non-telomere signals or interstitial telomere signals can indicate that the chromosome has undergone structural or quantitative rearrangement [56]. (AG3T3)3 signals were found around the centromeres in T. wallichiana, T. × media, T. yunnanensis, T. cuspidata, and T. wallichiana var. mairei, suggesting that these species may have undergone chromosome recombination. Studies have shown that there is more frequent gene loss and gain and genome-scale genomic rearrangement in gymnosperms [57]. The occurrence of this phenomenon may be related to the loss of reverse repeat sequences [1,58]. However, we did not lose sight of the fact that, due to the limited resolution of the FISH technique, no small gaps in the telomere signals due to chromosomal fusion were detected.
The 5S rDNA is often used to identify differences between species and differences in the ploidy of the same species. It is also used to conduct species identification and the phylogenetic identification of extra-specific and inter-specific taxa [27,59,60]. The 5S rDNA signal may be located on every euchromosome [61], and may occur in the middle, near the middle, or at the end of the chromosome [24]. The 5S rDNA signal patterns were found at the centromeres of all five species of Taxus, but the 5S rDNA signal was also found at the proximal end of the chromosomes of T. wallichiana, T. cuspidata, and T. yunnanensis. Such results indicate that the 5S rDNA repeats are moving at each site, which may also be related to the deletion of inverted repeats (IRS). The signal patterns on the chromosomes of the five species of Taxus are different, which we believe may be because the plant adapts to the environmental disturbance and changes the gene expression profile to adapt to the environmental change [62], resulting in more gene deletion and acquisition. However, genomic studies are hampered by the lack of reference genomes and transcriptome of non-model plants, so the genomic level of gymnosperms is still being explored [63].
The signal patterns on the chromosomes of the five species of Taxus are different, which we believe may be because the plant adapts to environmental disturbances and changes the gene expression profile to adapt to the environmental change [62], resulting in more gene deletion and acquisition. However, genomic studies are hampered by the lack of reference genomes and the transcriptome of non-model plants, so the genomic level of gymnosperms is still being explored [63]. Of course, the more plausible reasons need to be further studied. The Taxus plants classification is mainly based on morphological description and geographical distribution [11], but some species of Taxus have little difference in phenotypic characteristics [9], which leads to the classification of species of Taxus being controversial. Chromosome data are useful for plant classification [64]. Hao et al. [1] inferred from chloroplast intergenic spacer and nuclear coding DNA that T. wallichiana var. mairei and T. yunnanensis are more closely related than T. × media [1]. Liu et al. [65] studied the phylogenetic relationship of Taxus from the perspective of Internal Transcribed Spacer (ITS), but excluded several controversial species such as T. sumatrana and T. yunnanensis. The 5S rDNA signals, which vary in location and intensity, were useful for intraspecific and interspecific taxonomic identification and phylogenetic relationship characterization [27]. 5S rDNA and (AG3T3)3 signals were shown to be located at different locations in five species in Taxus. Identification using only two signals might be unscientific, but it will contribute in providing a classification reference in Taxus.
There is a phylogenetic framework for variation in 5S rDNA distribution because taxa closeness is associated with a similarity in the FISH signal pattern [66,67]. The 5S rDNA signals were used as the main differentiators and (AG3T3)3 signals were auxiliary to determine the genetic relationship. The phylogenetic relationships between T. cuspidate and T. × media and between T. wallichiana and T. yunnanensis were the closest. On the other hand, the T. wallichiana var. mairei species was phylogenetically distant. The experimental results were similar to those of Wang et al. [68]. The discrepancy with previous results may be due to the different methods used. In addition, different origins and hybridization conditions may affect the relationships between these species. However, the use of 5S rDNA main signals and (AG3T3)3 auxiliary signals to determine the genetic relationship might be unscientific, so it should therefore be certified by more signal probes. In order to further study the phylogenetic relationships among the five Taxus species, molecular data need to be combined.
The results of the oligonucleotide probes of five Taxus species were different, which may be helpful to draw oligonucleotide maps of Taxus plants in the future. If a specific oligonucleotide probe of Taxus is clear, it will be very convenient to identify Taxus. At the same time, we analysed the genetic relationship of five Taxus plants, which will be conducive to future cross breeding and use of heterosis.
5. Conclusions
Based on oligo-FISH, we found that the karyotype of the five Taxus species are different. This study provides chromosome characters that may be useful for karyotype comparisons with better pair identification within and between specific chromosome sets, as well as a physical map, contributing to the research on the cytogenetic composition of the Taxus genome.
Author Contributions
Z.H. and X.L. collected the materials and conducted the experiments. Z.H. and X.L. designed the oligo-probes, wrote the manuscript, provided funding, and supervised the study. Y.L. and W.Z. performed chromosome image analysis. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funding from the Key Research and Development Projects of Sichuan (No. 2018NZ0057) and the National Natural Science Foundation of China (No. 31500993).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data and materials are included in the form of graphs in this article.
Acknowledgments
The authors thank Zhou Yonghong for the support with laboratory equipment. We are also grateful to Wei Meng for growing the Taxus saplings.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hao, D.C.; Huang, B.; Yang, L. Phylogenetic relationships of the genus Taxus inferred from chloroplast intergenic spacer and nuclear coding DNA. Biol. Pharm. Bull. 2008, 31, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanović, M.; Ristić, M.; Popović, Z.; Matić, R.; Nikolić, B.; Vidaković, V.; Obratov-Petković, D.; Bojović, S. Chemical composition and interpopulation variability of essential oils of Taxus baccata L. from Serbia. Chem. Biodivers. 2016, 13, 943–953. [Google Scholar] [CrossRef]
- Krauze-Baranowska, M. Flavonoids from the genus Taxus. Z. Nat. C J. Biosci. 2004, 59, 43–47. [Google Scholar] [CrossRef]
- Kumar, S.; Mahdi, H.; Bryant, C.; Shah, J.P.; Garg, G.; Munkarah, A. Clinical trials and progress with paclitaxel in ovarian cancer. Int. J. Womens Health 2010, 2, 411–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, S.; Belani, C.P. Paclitaxel for non-small cell lung cancer. Expert Opin. Pharmacother. 2004, 5, 1771–1780. [Google Scholar] [CrossRef]
- Chen, T.K.; Wang, F.H. Embryogeny of Pseudotaxus chienii in relation to its systematic position. Acta Phytotaxon. Sin. 1978, 16, 1–10. Available online: http://www.plantsystematics.com/qikan/epaper/zhaiyao.asp?bsid=13297 (accessed on 2 September 2022).
- Hsu, C.Y.; Wu, C.S.; Chaw, S.M. Ancient nuclear plastid DNA in the yew family (Taxaceae). Gen. Biol. Evol. 2014, 6, 2111–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Laubenfels, D.J. Coniferales. In Flora Male Siana; Series I; Kluwer Academic: Dordrecht, The Netherlands, 1988; Volume 10, pp. 337–453. Available online: https://www.researchgate.net/publication/284054343_Coniferales (accessed on 2 September 2022).
- Zhou, Q.X.; Ge, S.; Gu, Z.J.; Yue, Z.S. Genetic variation and relationships within Taxus and between the genus and Pseudotaxus in China. Acta Phytotaxon. Sin. 1999, 36, 323. Available online: http://www.cnki.com.cn/Article/CJFDTotal-ZWFX804.005.htm (accessed on 2 September 2022).
- Fu, L.G.; Li, N.; Mill, R.R. Cephalotaxaceae. In Flora of China; Science Press: Beijing, China, 1999; pp. 89–96. [Google Scholar]
- Eckenwalder, J.E. Conifers of the World: The Complete Reference; Timber Press: Portland, Oregon, 2009; ISBN 9780881929744. [Google Scholar]
- Cheng, Y.C.; Nicolson, R.G.; Tripp, K.; Chaw, S.M. Phylogeny of Taxaceae and Cephalotaxaceae genera inferred from chloroplast matK gene and nuclear rDNA ITS region. Mol. Phylogenet. Evol. 2000, 14, 353–365. [Google Scholar] [CrossRef]
- Farjon, A. A Handbook of the World’s Conifers; Brill Academic Publishers: Leiden, The Netherlands, 2010; ISBN 9789004177185. [Google Scholar]
- Farjon, A. World Checklist and Bibliography of Conifers; Royal Botanical Gardens at Kew: London, UK, 1998; p. 316. ISBN 1900347547. [Google Scholar]
- Gu, Z.J.; Zhou, Q.X.; Yue, Z.K. Karyomorphological study on the genera Taxus and Pseudotaxus in China. Acta Bot. Yunnanica 1998, 20, 329–333. Available online: http://www.cnki.com.cn/Article/CJFDTotal-YOKE803.016.htm (accessed on 2 September 2022).
- Zhang, Y.Z.; Ma, J.; Yang, B.X.; Li, R.Y.; Zhu, W.; Sun, L.L.; Tian, J.K.; Zhang, L. The complete chloroplast genome sequence of Taxus chinensis var. mairei (Taxaceae): Loss of an inverted repeat region and comparative analysis with related species. Gene 2014, 540, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Kan, S.L.; Shen, T.T.; Gong, P.; Ran, J.H.; Wang, X.Q. The complete mitochondrial genome of Taxus uspidate (Taxaceae) eight protein-coding genes have transferred to the nuclear genome. BMC Evol. Biol. 2020, 20, 10. [Google Scholar] [CrossRef] [Green Version]
- Cusido, R.M.; Onrubia, M.; Sabater-Jara, A.B.; Moyano, E.; Bonfill, M.; Goossens, A.; Angeles Pedreno, M.; Palazon, J. A rational approach to improving the biotechnological production of taxanes in plant cell cultures of Taxus spp. Biotechnol. Adv. 2014, 32, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Nystedt, B.; Street, N.R.; Wetterbom, A.; Zuccolo, A.; Lin, Y.C.; Scofield, D.G.; Vezzi, F.; Delhomme, N.; Giacomello, S.; Alexeyenko, A.; et al. The Norway spruce genome sequence and conifer genome evolution. Nature 2013, 497, 579–584. [Google Scholar] [CrossRef] [Green Version]
- Dubreuil, M.; Riba, M.; Gonzalez-Martinez, S.C.; Vendramin, G.G.; Sebastiani, F.; Mayol, M. Genetic effects of chronic habitat fragmentation revisited: Strong genetic structure in a temperate tree, Taxus baccata (Taxaceae), with great dispersal capability. Am. J. Bot. 2010, 97, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasiri, J.; Naghavi, M.R.; Alizadeh, H.; Moghadam, M.R.F. Seasonal-based temporal changes fluctuate expression patterns of TXS, DBAT, BAPT and DBTNBT genes alongside production of associated taxanes in Taxus baccata. Plant Cell Rep. 2016, 35, 1103–1119. [Google Scholar] [CrossRef] [PubMed]
- Tomasino, M.P.; Gennaro, A.; Simeone, M.C.; Schirone, B.; Ceoloni, C. New insights into the Taxus baccata L. karyotype based on conventional and molecular cytogenetic analyses. Caryologia 2017, 70, 248–257. [Google Scholar] [CrossRef]
- Doležel, J. Plant Genome Diversity; Springer: Vienna, Austria, 2013; Volume 2, pp. 137–149. [Google Scholar]
- Luo, X.M.; Liu, J.C.; Zhao, A.J.; Chen, X.H.; Wan, W.L.; Chen, L. Karyotype analysis of Piptanthus concolor based on FISH with an oligonucleotide for rDNA 5S. Sci. Hortic. 2017, 226, 361–365. [Google Scholar] [CrossRef]
- Lan, Y.; Qu, L.W.; Xin, H.Y.; Gong, H.L.; Lei, J.J.; Xi, M.L. Physical mapping of rDNA and karyotype analysis in Tulipa sin-kiangensis and T. schrenkii. Sci. Hortic. 2018, 240, 638–644. [Google Scholar] [CrossRef]
- De Oliveira Bustamante, F.; do Nascimento, T.H.; Montenegro, C.; Dias, S.; do Vale Martins, L.; Braz, G.T.; Benko-Iseppon, A.M.; Jiang, J.; Pedrosa-Harand, A.; Brasileiro-Vidal, A.C. Oligo-FISH barcode in beans: A new chromosome identification system. Theor. Appl. Genet. 2021, 134, 3675–3686. [Google Scholar] [CrossRef]
- He, Z.J.; Zhang, W.; Luo, X.M.; Huan, J. Five Fabaceae karyotype and phylogenetic relationship analysis based on oligo-FISH for 5S rDNA and (AG3T3)3. Genes 2022, 13, 768. [Google Scholar] [CrossRef] [PubMed]
- Hizume, M.; Shibata, F.; Maruyama, Y.; Kondo, T. Cloning of DNA sequences localized on proximal fluorescent chromosome bands by microdissection in Pinus densiflora Sieb. and Zucc. Chromosoma 2001, 110, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.M.; Chen, J.Y. Physical map of FISH 5S rDNA and (AG3T3)3 signals displays Chimonanthus campanulatus R.H. Chang & C.S. Ding chromosomes, reproduces its metaphase dynamics and distinguishes its chromosomes. Genes 2019, 10, 904. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.H.; Cai, Z.X.; Xiang, S.Q.; Guo, Q.G.; Huang, W.; Liang, G.L. Karyotype analysis of diploid and spontaneously occurring tetraploid blood orange [Citrus sinensis (L.) Osbeck] using multicolor FISH with repetitive DNA sequences as probes. Front. Plant Sci. 2019, 10, 331. [Google Scholar] [CrossRef]
- Deng, H.H.; Xiang, S.Q.; Guo, Q.G.; Jin, W.W.; Liang, G.L. Molecular cytogenetic analysis of genome-specific repetitive elements in Citrus clementina Hort. Ex Tan. and its taxonomic implications. BMC Plant Biol. 2019, 19, 77. [Google Scholar] [CrossRef]
- Yi, K.U.; Kim, H.B.; Song, K.J. Karyotype diversity of Korean landrace mandarins by CMA banding pattern and rDNA loci. Sci. Hortic. 2018, 228, 26–32. [Google Scholar] [CrossRef]
- Doudrick, R.L.; Heslop-Harrison, J.; Nelson, C.D.; Schmidt, T.; Nance, W.L.; Schwarzacher, T. Karyotype of Slash pine (Pinus elliottii var. elliottii) using patterns of fluorescence in situ hybridization and fluorochrome banding. J. Hered. 1995, 86, 289–296. [Google Scholar] [CrossRef]
- Jacobs, M.D.; Gardner, R.C.; Murray, B.G. Cytological characterization of heterochromatin and rDNA in Pinus radiata and P. taeda. Plant Syst. Evol. 2000, 223, 71–79. [Google Scholar] [CrossRef]
- Vischi, M.; Jurman, I.; Bianchi, G.; Morgante, M. Karyotype of Norway spruce by multicolor FISH. Theor. Appl. Genet. 2003, 107, 591–597. [Google Scholar] [CrossRef]
- Fuchs, J.; Brandes, A.; Schubert, I. Telomere sequence localization and karyotype evolution in higher plants. Plant Syst. Evol. 1995, 196, 227–241. [Google Scholar] [CrossRef]
- Lubaretz, O.; Fuchs, J.; Ahne, R.; Meister, A.; Schubert, I. Karyotyping of three Pinaceae species via fluorescent in situ hybridization and computer-aided chromosome analysis. Theor. Appl. Genet. 1996, 92, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Doudrick, R.L.; Heslop-Harrison, J.S.; Schmidt, T. The contribution of short repeats of low sequence complexity to large conifer genomes. Theor. Appl. Genet. 2000, 100, 7–14. [Google Scholar] [CrossRef]
- Luo, X.M.; He, Z.J. Distribution of FISH oligo-5S rDNA and oligo-(AGGGTTT)3 in Hibiscus mutabilis L. Genome 2021, 64, 655–664. [Google Scholar] [CrossRef]
- Luo, X.M.; Liu, J.C.; He, Z.J. Oligo-FISH can identify chromosomes and distinguish Hippophaë rhamnoides L. Taxa. Genes 2022, 13, 195. [Google Scholar] [CrossRef]
- Qi, Z.X.; Zeng, H.; Li, X.L.; Chen, C.B.; Song, W.Q.; Chen, R.Y. The molecular characterization of maize B chromosome specific AFLPs. Cell Res. 2002, 12, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Hao, D.C.; Ge, G.B.; Xiao, P.G.; Zhang, Y.Y.; Yang, L. The first insight into the tissue specific Taxus transcriptome via Illumina second generation sequencing. PLoS ONE 2011, 6, e21220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.K.; Jin, Y.W.; Park, J.H.; Yoo, Y.M.; Hong, S.M.; Amir, R.; Yan, Z.; Kwon, E.; Elfick, A.; Tomlinson, S.; et al. Cultured cambial meristematic cells as a source of plant natural products. Nat. Biotechnol. 2010, 28, 1213–1217. [Google Scholar] [CrossRef]
- Wang, X.Q.; Ran, J.H. Evolution and biogeography of gymnosperms. Mol. Phylogenet. Evol. 2014, 75, 24–40. [Google Scholar] [CrossRef]
- Lu, Y.; Ran, J.H.; Guo, D.M.; Yang, Z.Y.; Wang, X.Q. Phylogeny and divergence times of gymnosperms inferred from single-copy nuclear genes. PLoS ONE 2014, 9, e107679. [Google Scholar] [CrossRef]
- Isah, T. Natural sources of taxol. J. Pharm. Res. Int. 2015, 6, 214–227. [Google Scholar] [CrossRef]
- Wang, B.Y. Reproductive Ecology of Taxus yunnanensis. Chinese Academy of Forestry. 2008. Available online: http://cdmd.cnki.com.cn/Article/CDMD-82201-2008136709.htm (accessed on 8 May 2022).
- Chen, L.; Cui, F.F.; Song, Y.; Zhu, Q.M.; Qiao, Y.G. Karyotype analysis of Taxus Media. Shanxi For. Sci. Technol. 2017, 46, 37–39. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-SXLK201703010.htm (accessed on 8 May 2022).
- Wang, S.H.; Qi, H.F.; Yuan, Y.J. Study on mitosis and chromosome stability of callus of Taxus cuspidate. Physiol. Mol. Biol. Plants 2000, 3, 263–266. Available online: http://www.cnki.com.cn/Article/CJFDTOTAL-ZWSI200003015.htm (accessed on 8 May 2022).
- Chen, K.Y. Karyotype analysis of Taxus cuspidate. Chin. Bull. Bot. 1996, 13, 46–47. Available online: https://www.chinbullbotany.com/CN/Y1996/V13/I/46 (accessed on 8 May 2022).
- Piednoël, M.; Aberer, A.J.; Schneeweiss, G.M.; Macas, J.; Novak, P.; Gundlach, H.; Temsch, E.M.; Renner, S.S. Next-generation sequencing reveals the impact of repetitive DNA in phylogenetically closely related genomes of Orobanchaceae. Mol. Biol. Evol. 2012, 29, 3601–3611. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, J.S.; Kim, H.; Nason, J.D.; Wing, R.A.; Wendel, J.F. Differential lineage-specific amplification of transposable elements is responsible for genome size variation in Gossypium. Genome Res. 2006, 16, 1252–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, B.G.; Friesen, N.; Heslop-Harrison, J.S. Molecular cytogenetic analysis of Podocarpus and comparison with other gymnosperm species. Ann. Bot. 2002, 89, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Majerová, E.; Mandáková, T.; Vu, G.T.; Fajkus, J.; Lysak, M.A.; Fojtová, M. Chromatin features of plant telomeric sequences at terminal vs. internal positions. Front. Plant Sci. 2014, 5, 593. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.C.; Luo, X.M. First report of bicolour FISH of Berberis diaphana and B. soulieana reveals interspecific differences and co-localization of (AGGGTTT)3 and rDNA 5S in B. diaphana. Hereditas 2019, 156, 13–21. [Google Scholar] [CrossRef]
- Luo, X.M.; Liu, J.C. Fluorescence in situ hybridization (FISH) analysis of the locations of the oligonucleotides 5S rDNA, (AGGGTTT)3, and (TTG)6 in three genera of Oleaceae and their phylogenetic framework. Genes 2019, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- Jansen, R.K.; Cai, Z.Q.; Raubeson, L.A.; Daniell, H.; Depamphilis, C.W.; Leebens-Mack, J.; Müller, K.F.; Guisinger-Bellian, M.; Haberle, R.C.; Hansen, A.K.; et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 19369–19374. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.F.; Li, Y.Q.; Yuan, X.L.; Hua, M.; Wang, Y.; Zhang, J. The complete chloroplast genome sequence of Taxus yunnanensis. Mitochondrial DNA B Resour. 2020, 5, 2756–2757. [Google Scholar] [CrossRef] [PubMed]
- Hasterok, R.; Wolny, E.; Hosiawa, M.; Kowalczyk, M.; Kulak-Ksiazczyk, S.; Ksiazczyk, T.; Heneen, W.K.; Maluszynska, J. Comparative analysis of rDNA distribution in chromosomes of various species of Brassicaceae. Ann. Bot. 2006, 97, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malinska, H.; Tate, J.A.; Matyasek, R.; Leitch, A.R.; Soltis, D.E.; Soltis, P.S.; Kovarik, A. A Similar patterns of rDNA evolution in synthetic and recently formed natural populations of Tragopogon (Asteraceae) allotetraploids. BMC Evol. Biol. 2010, 10, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.; Vargas, P.; Luceno, M.; Cuadrado, A. Evolution of Iris subgenus Xiphium based on chromosome numbers, FISH of nrDNA (5S, 45S) and trnL-trnF sequence analysis. Plant Syst. Evol. 2010, 289, 223–235. [Google Scholar] [CrossRef]
- Blödner, C.; Goebel, C.; Feussner, I.; Gatz, C.; Polle, A. Warm and cold parental reproductive environments affect seed properties, fitness, and cold responsiveness in Arabidopsis thaliana progenies. Plant Cell Environ. 2007, 30, 165–175. [Google Scholar] [CrossRef]
- Majeed, A.; Singh, A.; Sharma, R.K.; Jaitak, V.; Bhardwaj, P. Comprehensive temporal reprogramming ensures dynamicity of transcriptomic profile for adaptive response in Taxus contorta. Mol. Genet Genom. 2020, 295, 1401–1414. [Google Scholar] [CrossRef]
- Hu, J.Y.; Su, Z.X.; Yue, B.L.; Zhang, Q.Y. Progress of karyotype analysis method in plants research. J. Sichuan Teach. Coll. (Nat. Sci.) 2002, 23, 239–244. Available online: https://europepmc.org/article/CBA/377316 (accessed on 8 April 2022).
- Liu, J.; Moller, M.; Gao, L.M.; Zhang, D.Q.; Li, D.Z. DNA barcoding for the discrimination of Eurasian yews (Taxus L.; Taxaceae) and the discovery of cryptic species. Mol. Ecol. Resour. 2001, 11, 89–100. [Google Scholar] [CrossRef]
- Cai, Q.; Zhang, D.M.; Liu, Z.L.; Wang, X.R. Chromosomal localization of 5S and 18S rDNA in five species of Subgenus strobus and their implications for genome evolution of Pinus. Ann. Bot. 2006, 97, 715–722. [Google Scholar] [CrossRef]
- Wang, G.R.; Li, B.L.; Gao, S.L. RAPD Analysis and chromosome identification on several Taxus L. in China. Pharm. Biotechnol. 2005, 2, 81–84. Available online: http://en.cnki.com.cn/Article_en/CJFDTOTAL-YWSW200502003.htm (accessed on 8 May 2022).
- Wang, Y.F.; Shi, Q.W.; Dong, M.; Kiyota, H.; Gu, Y.C.; Cong, B. Natural Taxanes: Developments Since 1828. Chem. Rev. 2011, 111, 7652–7709. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphological comparison of the leaves and bud. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei.
Figure 1.
Morphological comparison of the leaves and bud. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei.

Figure 2.
Visualization of the mitotic metaphase chromosomes of T. wallichiana (A), T. × media (B), T. yunnanensis (C), T. cuspidate (D), and T. wallichiana var. mairei (E) after fluorescence in situ hybridization (FISH). The first probe of (AG3T3)3 was labelled with 6-carboxyfluorescein (FAM) (green); the second probe of 5S rDNA was labelled with 6-carboxytetramethylrhodamine (TAMRA) (red). Chromosomes probed with 5′-TAMRA-labelled 5S rDNA (red fluorescence, arrow) are shown in (A–E). The concentration of all the probes was 10 µM. All the chromosomes were counterstained with 4,6-diamidino-2-phenylindole (DAPI) (blue). Scale bar = 5 µm.
Figure 2.
Visualization of the mitotic metaphase chromosomes of T. wallichiana (A), T. × media (B), T. yunnanensis (C), T. cuspidate (D), and T. wallichiana var. mairei (E) after fluorescence in situ hybridization (FISH). The first probe of (AG3T3)3 was labelled with 6-carboxyfluorescein (FAM) (green); the second probe of 5S rDNA was labelled with 6-carboxytetramethylrhodamine (TAMRA) (red). Chromosomes probed with 5′-TAMRA-labelled 5S rDNA (red fluorescence, arrow) are shown in (A–E). The concentration of all the probes was 10 µM. All the chromosomes were counterstained with 4,6-diamidino-2-phenylindole (DAPI) (blue). Scale bar = 5 µm.

Figure 3.
Mitotic chromosomes of five Taxus species rearranged from Figure 1. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei. Each chromosome is numbered according to its length, from the longest to the shortest chromosome. Scale bars range from 1 to 15 μm.
Figure 3.
Mitotic chromosomes of five Taxus species rearranged from Figure 1. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei. Each chromosome is numbered according to its length, from the longest to the shortest chromosome. Scale bars range from 1 to 15 μm.

Figure 4.
Karyotype idiograms of five Taxus species. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei. The x-axis indicates the chromosome number, whereas the y-axis indicates the relative chromosome length.
Figure 4.
Karyotype idiograms of five Taxus species. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei. The x-axis indicates the chromosome number, whereas the y-axis indicates the relative chromosome length.

Figure 5.
Signal distribution of the five Taxus species. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei. The idiograms were constructed based on the signal patterns of the chromosomes mentioned above and the chromosomes in Figure 1. The numbers at the bottom represent the number of chromosomes.
Figure 5.
Signal distribution of the five Taxus species. (A) T. wallichiana, (B) T. × media, (C) T. yunnanensis, (D) T. cuspidate, and (E) T. wallichiana var. mairei. The idiograms were constructed based on the signal patterns of the chromosomes mentioned above and the chromosomes in Figure 1. The numbers at the bottom represent the number of chromosomes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Collection of seedling resources.
| No. | Species | Location | Tissue |
|---|---|---|---|
| 1 | Taxus wallichiana | Xichang City, Sichuan Province, China | Seedling |
| 2 | Taxus × media | Ya’an City, Sichuan Province, China | Seedling |
| 3 | Taxus yunnanensis | Ya’an City, Sichuan Province, China | Seedling |
| 4 | Taxus cuspidata | Suqian City, Jiangsu Province, China | Seedling |
| 5 | Taxus wallichiana var. mairei | Ya’an City, Sichuan Province, China | Seedling |
Table 2.
Karyotype formulas of five species of Taxus.
| Species | Karyotype | Karyotype Asymmetry Index | Arm Ratio |
|---|---|---|---|
| T. wallichiana | 2n = 2x = 24m | 55.56% | 3.0087 |
| T. × media | 2n = 2x = 24m | 55.09% | 3.4198 |
| T. yunnanensis | 2n = 2x = 24m | 55.56% | 2.6402 |
| T. cuspidate | 2n = 2x = 24m | 54.67% | 3.0135 |
| T. wallichiana var. mairei | 2n = 2x = 22 m + 2sm | 54.33% | 2.8716 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
He, Z.; Luo, X.; Lei, Y.; Zhang, W. Five Species of Taxus Karyotype Based on Oligo-FISH for 5S rDNA and (AG3T3)3. Genes 2022, 13, 2209. https://doi.org/10.3390/genes13122209
AMA Style
He Z, Luo X, Lei Y, Zhang W. Five Species of Taxus Karyotype Based on Oligo-FISH for 5S rDNA and (AG3T3)3. Genes. 2022; 13(12):2209. https://doi.org/10.3390/genes13122209
Chicago/Turabian StyleHe, Zhoujian, Xiaomei Luo, Yuting Lei, and Wei Zhang. 2022. "Five Species of Taxus Karyotype Based on Oligo-FISH for 5S rDNA and (AG3T3)3" Genes 13, no. 12: 2209. https://doi.org/10.3390/genes13122209
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.