1. Introduction
Climate change is a global crisis and challenge, which has and will pose nonnegligible impacts on the natural ecosystem and human survival and development [
1]. As indicated in ICPP AR6, over the past 100 years, the global temperature elevated by 1.1 °C relative to that during the preindustrial period [
2]. Climate is the most vital environmental factor determining tree species distribution [
3], while climate change has resulted in tremendous changes of the suitable habitats for plants and also influenced the local original ecosystem and the ecological service function [
4,
5]. It has been indicated that plants migrate to the higher altitude or higher latitude areas [
6], and two thirds of the spring phenology has been advanced [
7]. Under the background of global warming, extreme weathers occur frequently, severely impairing tree species [
8]. The world biodiversity is disappearing at an unprecedented rate [
9], with approximately 100 species becoming extinct every day [
10], and climate change has become the major threat to biodiversity and ecosystems [
11,
12,
13,
14,
15,
16,
17,
18].
Relict plants are species that were once widely distributed in the Tertiary Arctic but are currently only distributed in East Asia, North America, and southwestern Europe [
19]. Honored as the living fossil plants, the tertiary relict plants have aroused extensive attention due to their ancient origin, special morphology, and significant research value [
20,
21], also becoming the hot topic in the field of biodiversity protection [
22,
23,
24]. During the Neogene period and the Quaternary period, due to global climate change, these species that were once widely distributed in the Northern Hemisphere began to migrate to the warm and humid areas in North America, East Asia, and Southwest Eurasia [
25,
26]. The flora in East Asia has the most abundant species diversity, with an ancient and complete succession sequence, which can provide vital information for the lost historical records and is also an important area to explore in terms of plant origin and evolution in the northern hemisphere [
26,
27]. At present, numerous relict plants (
Appendix A) are listed as endangered species, including
Shaniodendron subaequale [
28],
Liriodendron chinense [
29],
Taxus wallichiana [
30], and
Ginkgo biloba [
31]. For relict plants, climate change will not only lead to the degradation or loss of their habitats [
29,
32] but also block gene communications between populations, affecting their genetic diversity and adaptability to the environment [
33,
34]. Therefore, it is of great significance to investigate the adaptive mechanisms of relict plants to the global climate change for their protection and sustainable development.
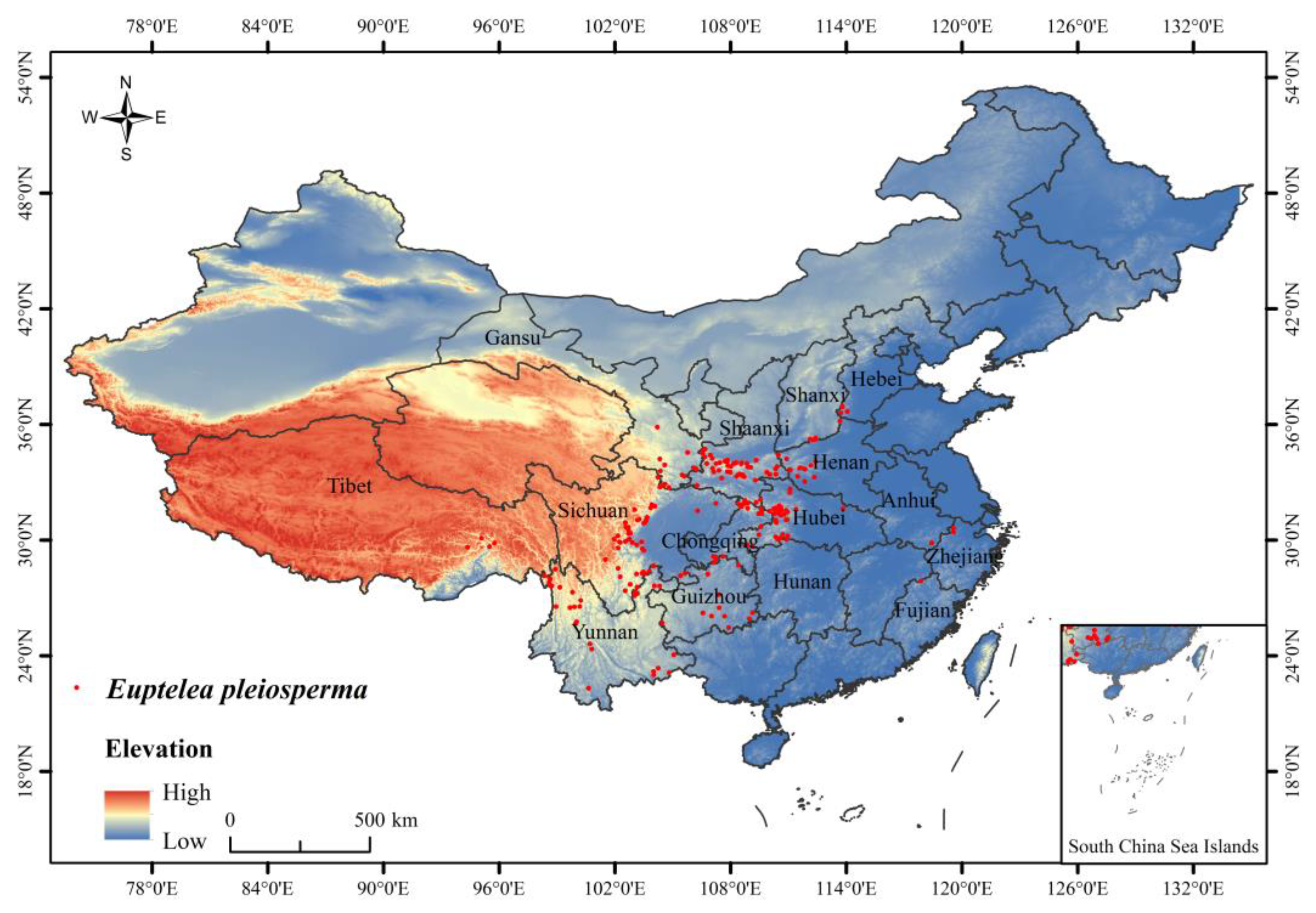
Euptelea pleiosperma Hook. f. et Thoms is a small deciduous tree belonging to Euptelea of Eupteleaceae [
35]. During the Late Miocene, global cooling resulted in
E. pleiosperma populations in China gradually migrating to the lower latitude area [
36]. They sought suitable habitats while migrating along the altitude and used mountains as shelters [
19]. The survey shows that
E. pleiosperma is distributed in Zhejiang, Hunan, Hubei, Guizhou, Yunnan, Shanxi, Hebei, and Henan in China [
37]. Eupteleaceae is a monotypic family endemic to East Asia, including one genus: Euptelea. There are only two species of this genus: one is distributed in China and India, and the other is originated in Japan. The distribution and number of
E. pleiosperma are extremely limited. If
E. pleiosperma becomes extinct in China, it will generate a certain impact on its biodiversity [
35]. In terms of phylogeny, this species has an isolated status, possessing several relatively primitive features and having few closely related species.
E. pleiosperma belongs to the tertiary relict plant group and is also the typical characteristic species as the floristic component of East Asia. At present, studies on
E. pleiosperma mainly concentrate on seed traits [
38], genetic diversity [
39,
40], and community structure [
35], while studies regarding the response to climate and the estimation of the potential distribution are still lacking. However, with the aggravation of global climate change and excessive disturbance of human activities, the natural habitat for
E. pleiosperma is severely fragmented, its distribution range is shrinking, and its population quantity is decreasing and even on the edge of extinction; it is currently listed in the Red Book of Chinese Plants as a Grade III protected plant in China [
22]. Therefore, determining the potential geographical distribution of
E. pleiosperma and estimating the influence of future climate change on its suitable habitat can provide effective information for protecting population resources.
Species distribution models (SDMs), also referred to as niche models, combine the observed species distribution data with the corresponding environmental variables in order to infer the ecological needs of species, which can thus estimate their potential distribution [
41]. In addition, there is a growing trend to utilize SDMs to investigate the influence of climate on species distribution [
42,
43]. Scholars have developed multiple models based on different algorithms [
44,
45], which are extensively applied in numerous fields including species protection, natural environment monitoring, evaluation of ecological situation, protection against invasive species, and estimation of the risk of infection [
44,
46,
47,
48,
49]. This study used three models (BIOCLIM, DOMAIN, and MaxEnt) to estimate the suitable habitats of species [
50,
51]. The BIOCLIM model is a framework niche model based on bioclimatic data, which can extract various environmental data from the known distribution areas to form fixed and complex rectangle packing. If the values of regional environmental variables are within the range of the rectangle packing, then it is shown that this region is suitable for species distribution [
52]. The DOMAIN model calculates the Gower distance between points to determine the maximum similarity degree of environmental factors between the known distribution area and the estimated area, which can therefore judge the species distribution range [
53]. MaxEnt is a machine-learning algorithm based on the principle of maximum entropy, which can simulate the species niches using data related to the environmental variables and habitat suitability; in addition, the distribution probability can be estimated by the maximum entropy distribution restricted by the environmental variables [
54].
Studies on the potential distribution of species have begun to use niche models, and each niche model exhibits specific preferences. Different results may be obtained from the same species distribution data when they are applied in different models. In order to maximally lower the simulation deviation caused by the application of a single model, this study integrated the BIOCLIM, DOMAIN, and MaxEnt models and selected the optimal one to estimate the future potential suitable areas of E. pleiosperma in China. This study aimed to (i) investigate the estimation accuracy of three niche models and select the optimal model for simulation; (ii) investigate the major environmental factors affecting E. pleiosperma distribution; and (iii) summarize the potential suitable habitat patterns of E. pleiosperma and the dynamic variation rules under different climate scenarios.
3. Results
3.1. Model Accuracy Evaluation
The mean AUC values and Kappa values of the BIOCLIM, DOMAIN, and MaxEnt models from ten-fold simulations are displayed in
Figure 2. The mean AUC values of the three models were 0.929, 0.898, and 0.788, respectively, higher than 0.7. This represents an excellent estimation accuracy. At the same time, the standard deviations were in the order of BIOCLIM (0.003) < MaxEnt (0.005) < DOMAIN (0.009). The Kappa values of the three models were 0.772, 0.627, and 0.576, respectively, higher than 0.55, suggesting good consistency test results. The standard deviations of the Kappa value were in the order of BIOCLIM (0.009) < MaxEnt (0.01) < DOMAIN (0.019). The AUC values of the three models were higher than their Kappa values. Furthermore, evaluation results by the two methods both revealed that BIOCLIM had the highest values, followed by DOMAIN, while the MaxEnt model exhibited the lowest values, indicating that the AUC values were partially associated with the Kappa values. The smaller standard deviations indicated a lower sample dispersion degree and a lower influence on the estimation results. In general, the MaxEnt model achieved an average accuracy, whereas the DOMAIN model had an excellent accuracy. The BIOCLIM model showed a good accuracy, and all of them accurately reflected the potential distribution of
E. pleiosperma. The BIOCLIM model had the highest scores of both evaluation indexes, the highest accuracy and the lowest standard deviation; thus, it was considered to be the optimal model.
3.2. Major Influencing Environmental Factors
The BIOCLIM and DOMAIN models could not explore the contributions of environmental factors to the estimation of distribution. If the environmental variables affecting the
E. pleiosperma distribution are analyzed, it is essential to estimate the combination of single variables or multiple variables, which takes a long time and is difficult to perform operationally on a large scale [
68]. The “knife-edge” function of the MaxEnt model can measure the importance of environmental variables and output the results directly when making the estimation. In addition, although the MaxEnt model shows a lower accuracy evaluation value than the other two models, which may have a bias in estimating the contribution of environmental factors, its accuracy is still within the reference range, and the results have reference significance. Therefore, based on the output information of the MaxEnt model, the major environmental factors affecting the potential distribution of
E. pleiosperma were obtained.
As presented in
Table 4, among the 11 environmental variables, bio6, bio12, and alt ranked the top three places, with contribution rates of 37.8%, 31.9%, and 13.5%, respectively, significantly higher than those of the other environmental factors. Moreover, in the jackknife test, the “without variable”, “with only variable”, and “with all variable” patterns were employed to explore the regularized training gain, also displaying the importance of the 11 environmental variables to the current species distribution.
Figure 3 shows that the top three factors with the highest training model scores were bio6, bio12, and bio2, respectively. Considering the contribution rates and importance, it was determined that the environmental variables which dominated the potential distribution areas of
E. pleiosperma included temperature (bio6, bio2), precipitation (bio12), and terrain (alt).
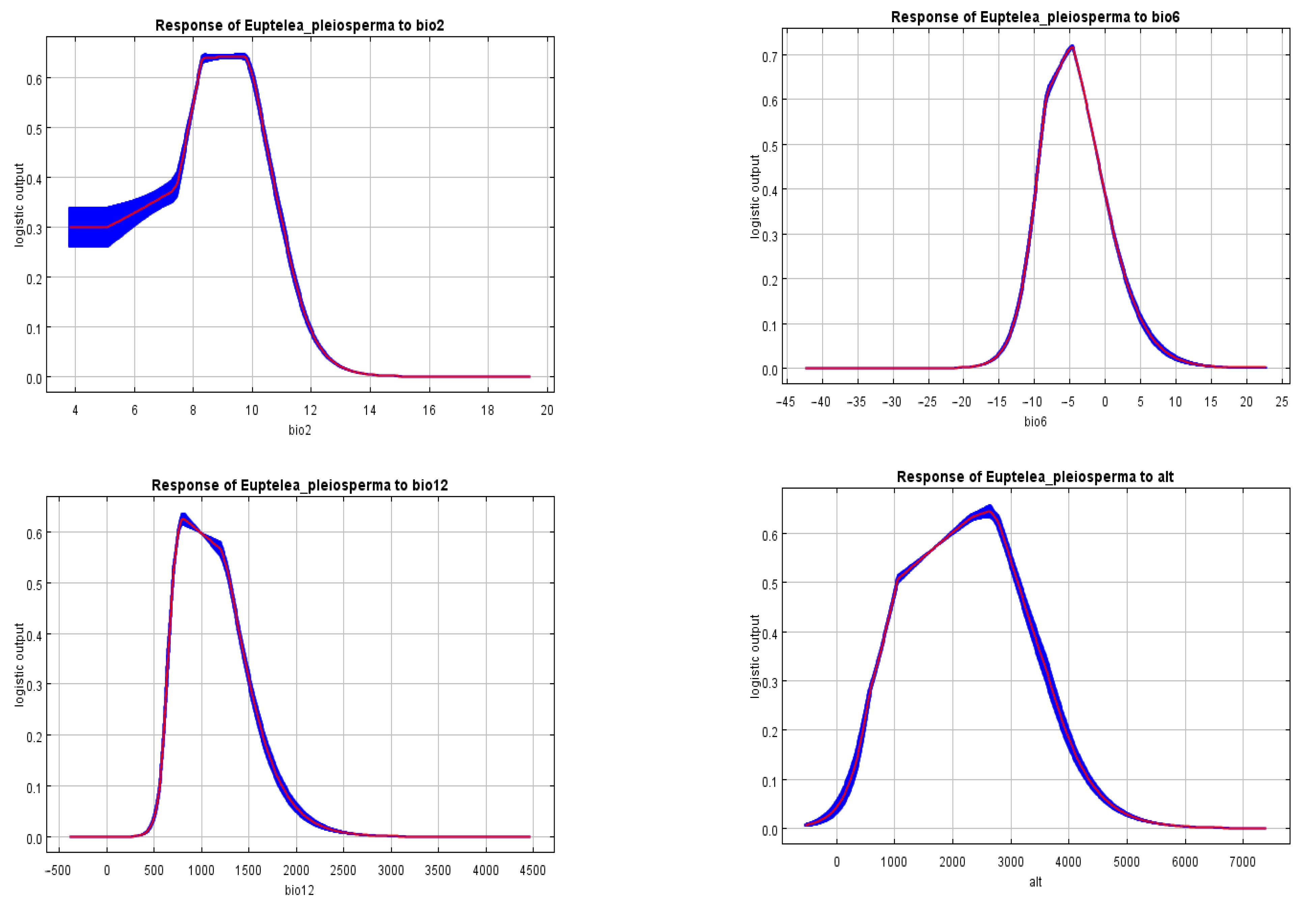
To intuitively understand the influencing mechanism of every factor on the suitable habitat of
E. pleiosperma, the MaxEnt model was used to plot the single-factor response curves, aiming to reflect the influence features of major environmental factors on the suitable habitat distribution probability and the species tolerance to the environment (
Figure 4). For the four environmental factors, their suitable habitat distribution probability exhibited the trend of first increasing and then decreasing. The peaks displayed the optimal distribution values. A higher suitable habitat distribution probability was maintained within bio2 values of 8.5–9.8 °C. The probability rapidly declined beyond this range. When the range was >12 °C, the suitable habitat distribution probability was lower than 0.08, indicating that the area was no longer suitable for the survival of
E. pleiosperma. Moreover, in terms of bio6, the suitable habitat distribution range was −13.4–6.1 °C, and the maximal probability value reached 0.718 at −4.7 °C. The suitable habitat range of bio12 was 545.8–1908.6 mm, and that of the highly suitable area was 691.8–1309.9 mm. The highest suitable habitat probability of 0.625 was found at 803.7 mm. In terms of altitude, the suitable habitat range was 175.9–4459.0 m, which was wider than the other environmental factors. The optimal suitable distribution range was 1026.2–3139.9 m, with the highest suitable distribution probability of 0.645 at the altitude of 2631.4 m.
3.3. Potential Suitable Habitats under Current Climates and Model Selection
The potential distribution estimation results of
E. pleiosperma in China based on the three models are displayed in
Figure 5 and
Table 5. According to the MaxEnt model results, the suitable habitat range was from 22° N~41° N to 79° E~125° E, and the total distribution area occupied 15.24% of the total land area, mainly exhibiting a zonal distribution in central and southwestern China, with sparse distribution in southeastern China. The highly suitable habitats were found in Shaanxi, Sichuan, Gansu, Tibet, Yunnan, Shanxi, Chongqing, Guizhou, Henan, Hubei, Anhui, and Shandong, with the area reaching 262.89 × 10
3 km
2. Using the BIOCLIM model, the suitable habitat range was from 24° N~37° N to 87° E~119° E, particularly from 24° N~37° N to 93° E~114° E. Compared with the MaxEnt model, the suitable distribution area estimated by the BIOCLIM model showed a lost trend, with an area of 804.54 × 10
3 km
2 occupying 8.35%. The highly suitable area occupied 0.67%, mainly distributed in Shaanxi, Sichuan, Gansu, Chongqing, Yunnan, Guizhou, Henan, and Hubei. The high-, medium-, and low-suitability habitat areas estimated by the MaxEnt and BIOCLIM models increased in succession. Using the BIOCLIM model, the areas of suitable habitats at various grades were lower than those estimated by the MaxEnt model, and the high suitability distribution area exhibited a narrow zonal distribution. The current suitable habitats of
E. pleiosperma estimated by the DOMAIN model are almost all over the country, which is extremely different from the other two models. In addition, Wang et al. [
50] and Duan et al. [
69] also obtained similar results, which may be a result of the false behavior of the model. This model was discarded due to the great deviation.
Although the potential geographical distribution regions and areas estimated by the MaxEnt and BIOCLIM models were different, the results were quite similar. The suitable areas were largely overlapping, especially for the highly suitable area range. The BIOCLIM model achieved the highest simulation accuracy, with its estimated current suitable area being located in the subtropical area in China. Basically, the results were consistent with the actual E. pleiosperma distribution area, suggesting that the BIOCLIM simulation results were relatively accurate and reliable. Therefore, the BIOCLIM model was selected to estimate the potential distribution of E. pleiosperma under the future climate scenario.
3.4. Estimation of the Potential Distribution of E. pleiosperma under the Future Climate Scenario
The potential suitable area distribution results estimated based on the BIOCLIM model under different emission scenarios (SSP126, SSP370, and SSP585) in 2041–2060 and 2081–2100 are displayed in
Figure 6 and
Table 6. The distribution of various suitable area grades in the future potential distribution area varied to varying degrees. Under different climate scenarios, the future suitable area still presented zonal distribution, accompanied by severe habitat fragmentation, and the high suitable area remained narrow. Under the SSP126 scenario, the total suitable area will increase from 8.36% to 8.70% and later decrease to 8.60%. The highly suitable habitat area will decrease from 0.67% to 0.60% and later increase to 0.69%. Under the SSP370 emission scenario, the total suitable habitat area will increase from 8.36% to 8.43% and subsequently decrease to 8.14%, while the highly suitable habitat area will increase from 0.67% to 0.72% and later decrease to 0.61%. Under the SSP585 scenario, the total suitable habitat area will elevate from 8.36% to 8.84% and then decrease to 5.75%. The highly suitable habitat area will increase from 0.67% to 0.79% and later decrease to 0.41%. Relative to the current scenario, the total suitable area under the future SSP126 scenario will finally increase to 0.25%, that under the SSP370 emission scenario will decrease by 0.21%, and that under the SSP585 emission scenario will decrease by 2.6%.
Under the three emission concentrations, the total suitable habitat area exhibited first a gain and then a loss trend from the current to the future scenarios. Apart from the SSP126 scenario, the variation trend in the highly suitable habitat area will be basically consistent with that of the total suitable habitat area. Under the SSP585 highest emission concentration scenario, E. pleiosperma will obtain the greatest highly suitable habitat area and total suitable habitat area in 2041–2060, compared to the smallest highly suitable habitat area and total suitable habitat area in 2081–2100. The low-suitability areas will account for a large proportion of the total suitable area, followed by medium-suitability areas, and high-suitability areas will occupy the lowest proportion.
3.5. Dynamic Variation in Potential Suitable Area under the Future Climate Scenarios
Based on the results estimated by the BIOCLIM model, the edge and central area in the current potential suitable habitat were greatly susceptible to climate change, exhibiting gain and loss to varying degrees. The change areas were sparsely distributed and fragmented (
Figure 7,
Table 7). Compared with the current potential distribution area, the gain and loss areas of the suitable habitat under the SSP126 scenario in 2041–2060 will be 107.25 × 10
3 km
2 and 70.95 × 10
3 km
2, respectively. To be specific, the gain areas will be concentrated in Sichuan, Shanxi, Henan, Hunan, and Yunnan, while the lost areas will be concentrated in Sichuan, Fujian, Gansu, and Ningxia. Under the SSP370 scenario, the gain area of suitable habitat will be 70.90 × 10
3 km
2, whereas the lost areas are still concentrated in the Sichuan area, with the total lost area of 61.79 × 10
3 km
2. Under the SSP585 scenario, the lost area of suitable habitat will be 60.95 × 10
3 km
2. The maximum gain area will be obtained (112.29 × 10
3 km
2), mainly in provinces including Henan, Shanxi, Shaanxi, Guangdong, and Hunan.
Compared with 2041–2060, more significant fluctuations will be observed in suitable habitats in 2081–2100. Under the SSP126 scenario, the gain areas will occur in the Gansu, Shaanxi, Tibet, Sichuan, Jiangxi, and Fujian areas, with a total area of 95.67 × 103 km2. Meanwhile, the lost areas will be highly similar to the gain areas from the current scenario to 2041–2060, with a total lost area of 104.61 × 103 km2. Under the SSP370 scenario, the gain and lost areas of suitable habitats will be close, mainly in Yunnan, Fujian, and Jiangxi. Under the SSP585 scenario, the suitable habitats will present a lost trend on the whole. The suitable habitats in Yunnan, Sichuan, Chongqing, Gansu, Shaanxi, Hunan, and Guangxi will be substantially decreased, the habitat degradation will be aggravated, and the total lost area will reach 401.74 × 103 km2, ranking top among the lost areas of suitable habitats among all the paths. In addition, this value is increased by 321.28 × 103 km2 compared with the gain area. Particularly, the suitable areas in the east of Tibet at the south of Gansu will increase significantly.
The gain area of suitable habitats will be greater than the lost area under the three emission scenarios in 2041–2060, indicating the increased area of suitable habitats during this future period. On the contrary, the lost area will be greater than the gain area in 2081–2100, indicating the gradually decreased area of suitable habitats during this future period. Moreover, the area of stable regions from the current to future climate scenarios will change significantly. Under the SSP585 emission scenario, the highest stable habitat area will be the highest (802.14 × 103 km2) in 2041–2060, while the lowest stable habitat area (512.69 × 103 km2) will be observed in 2081–2100.
3.6. Changes in Suitable Habitat Centroid under Different Climatic Scenarios
The centroid location can be calculated according to the potential suitable area of
E. pleiosperma under different climate scenarios estimated by the BIOCLIM model. Thus, the migration trajectory and trend of suitable habitats can be determined. As displayed in
Figure 8, under the SSP126 scenario, the centroid coordinates of the current suitable habitat are (29.30° N, 105.57° E), which will migrate to (29.30° N, 105.86° E) in 2041–2060, and to (29.21° N, 106.07° E) in 2081–2100. Under the SSP370 scenario, the centroid will migrate from (29.30° N, 105.57° E) to (29.34° N, 105.69° E) and later to (29.34° N, 105.86° E). Under the SSP585 scenario, the centroid will migrate from (29.30° N, 105.57° E) to (29.21° N, 105.89° E) and then to (29.71° N, 104.69° E).
In the future scenarios, the centroid of E. pleiosperma will mostly be distributed in the southwest of Chongqing City. However, under the SSP585 scenario in 2081–2100, the centroid will suddenly migrate to the northwest and finally to the southeast of Sichuan. The moving trajectory of the centroid also varies: migrating southeastward under the SSP126 scenario, northeastward under the SSP370 scenario, and northwestward under the SSP585 scenario. The centroid of the future suitable habitat will be evenly distributed in the southwest of China. Under the SSP370 medium-emission and SSP585 high-emission scenarios, the potential distribution areas may migrate to the higher northern latitude area, whereas they show a trend to the lower northern latitude areas under the SSP126 low-emission scenario.
5. Conclusions
To conclude, this study selected the optimal model from the three niche models (MaxEnt, BIOCLIM, and DOMAIN) to estimate the potential suitable habitat area of E. pleiosperma and to explore the pattern dynamics under climate change conditions. Our results suggested that the BIOCLIM model was the optimal model to estimate E. pleiosperma distribution and that temperature was the most critical factor restricting the geographical distribution pattern of E. pleiosperma, followed by precipitation and altitude. However, more research is needed to determine the specific contribution of each factor in order to determine the influencing factors more accurately, especially in methods using multiple SDM-model combinations. Under the current climate scenario, the potential suitable areas were mainly distributed in the central and southwestern areas. Under the future SSP126 scenario, the suitable habitat area showed expansion, and those under the other two emission scenarios exhibited shrinkage, with more fierce variation in the suitable habitat under the high-emission scenario. Under the medium- and high-emission scenarios, the potential suitable area for E. pleiosperma will migrate northward; under the low-emission scenario, the area will migrate southward. In the current high-suitability areas, the establishment of nature reserves and seed resource banks and the protection of wild populations can be carried out. Under the future climate scenarios, artificial guidance is recommended to ensure the smooth migration of E. pleiosperma to the expansion areas. Moreover, our research results can provide a reference for the protection and management of E. pleiosperma, which is of great benefit for sustainable development.
In future research work, the following four points need to be focused on: (i) the use of climate model groups composed of multiple climate models and improving data accuracy; (ii) the combination with other methods to screen the influencing variables, such as a random forest framework trained by the Boruta algorithm (B-RF); (iii) the incorporation of other influencing factors into the model; and (iv) further analysis of the specific response of individual trees or regional tree subpopulations to the environment and climate.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







