Allometric Model of Crown Length for Pinus sylvestris L. Stands in South-Western Poland


1
Institute of Biology, Faculty of Natural and Technical Sciences, University of Opole, Kominka 6A, 45-032 Opole, Poland
2
Institute of Environmental Engineering and Biotechnology, Faculty of Natural and Technical Sciences, University of Opole, Kominka 6A, 45-032 Opole, Poland
*
Author to whom correspondence should be addressed.
Forests 2023, 14(9), 1779; https://doi.org/10.3390/f14091779
Submission received: 12 August 2023
/
Revised: 28 August 2023
/
Accepted: 30 August 2023
/
Published: 31 August 2023
(This article belongs to the Section Forest Ecology and Management)
Abstract
:The growth of a tree depends on the size, shape, and functioning of the crown. The length of the crown is a somewhat subjective value because the base of the crown is often difficult to determine. The aim of this study was to develop an allometric model to calculate the crown length of Pinus sylvestris L., which might serve as an alternative to the current equations used especially for stands of variable density. The model used three predictive variables, i.e., diameter at breast height, tree height, and density. The developed crown length model showed high compatibility with empirical data within the studied stands differing in diameter at breast height, height, age, biosocial position, and, above all, density (SD = 1.786). The correlation coefficient between the empirical crown length for the stand () and the calculated model was r = 0.974, with a discrepancy of (±) 3.17%. The derived crown length model can be one of the components used to estimate the mass of needles or leaf area index (LAI) and, consequently, the amount of transpiration or the amount of carbon dioxide bound, which is crucial in the context of climate change.
1. Introduction
From an economic and environmental point of view, Scots pine (Pinus sylvestris L.) is one of the most important forest tree species, and it is the second largest in terms of its range among all coniferous species of the northern hemisphere [1,2]. In Poland, it is the dominant species that currently makes up about 58% of the forest species composition. This is due to the habitats typical for pine, i.e., coniferous and mixed coniferous forests, which cover about 50% of the forest area [3].
The size, structure, and shape of tree crowns determine the scope and efficiency of physiological processes, mainly photosynthesis, respiration, and transpiration, determining the growth and development of the tree. Estimates of crown size are often used to infer the tree’s vigour [4,5,6,7,8], basal area increment [8,9], production efficiency [10,11], and the quality and value of the wood [12,13,14,15,16]. In addition, knowledge of the size and shape of crowns is essential to predict the mechanical stability of trees [17] and their susceptibility to wind and snow damage [18,19,20,21], as well as vulnerability to fire [22,23,24]. It also helps to visualise the structure of trees and stands [18]. Furthermore, the crown length is used in allometric biomass models of biomass components, such as live and dead branches, stem bark, and leaves [25].
Burger [26,27,28,29,30,31] carried out extensive research on the structure and shape of tree crowns of various species. His research contributed to the development of crown models of such species as spruce, pine, or beech. In Poland, research on the relationship between crown size and tree growth in pine stands was carried out by Lemke [32,33,34]. Many works have focused on the relationship between the size of the crown and other features of the tree [35,36,37,38]. Diameter at breast height (DBH) is the most commonly used independent variable in crown width modelling, since the relationship between DBH and the crown width has been well established in the literature [39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54].
The length of the tree crown is of great importance in the methodology of breeding research [8,55,56,57,58]. This is because both the growth of the tree and the quality of the trunk depend on the length of its crown [59,60]. The length of the crown is difficult to determine if its shape is irregular. In forestry practice, it is measured from the top to the lowest green branch, maintaining continuity with the crown of the tree. Grochowski [61] considered the length of the crown to be the length from the top of the stem to the base of the first live branch, “maintaining continuity with the higher part of the crown”. If the crown is regular, that is, properly built, then its length is the distance from the treetop to the lowest base of the whorl of live branches.
Direct crown length measurements can be difficult to perform, especially in dense stands. Therefore, research works have begun to focus on the development of models for the crown length or relative crown length (i.e., the ratio of crown length to tree height). Numerous authors [62,63] have used simple geometric shapes for modelling. Other scientists began to use more flexible models to represent crown shapes [64,65,66,67,68]. Baldwin and Peterson [69] developed a series of equations to predict the outer and inner crown profile of Pinus taeda L., whereas Doruska and Mays [70], in an attempt to improve the crown profile estimate for this species, used nonparametric regression with only one predictive variable. The modelling was based on relationships between the tree and stand features [8,71,72,73]. Crown size predictions are usually based on allometric relationships with easy-to-measure tree trunk dimensions, such as tree height (H) and diameter at breast height (DBH). The use of these two biometric parameters for modelling seems justified because the size of the foliage determines the growth of trees in terms of their height and thickness [8,60,69,74,75,76]. In the case of stands growing at a low density (where there is no tree crown closure) or overpopulated stands, the use of these two parameters in allometric equations (H; DBH) might be insufficient to accurately determine the size of the crown [35,76,77]. Such equations may tend to overestimate the crown size in compact stands and underestimate it in open stands [78]. In such cases, it seems reasonable to add parameters related to competition, for example, taking density into account [35,41,54,79,80,81].
The aim of our research was to develop an equation to calculate the crown length (), which could serve as an alternative to current allometric equations, which are used especially in stands of varying density. It was hypothesised that of the three parameters, i.e., height, breast height, and density, the latter would have the greatest influence on crown length in pine stands. The following principal criteria were used to develop the model: (i) the model should account for the effect of density on the crown size; (ii) easy-to-measure biometric parameters of the tree, i.e., the height () and the diameter at breast height (—DBH) should be used to develop the equation; (iii) the equation should be capable of providing an accurate calculation of the crown length for Scots pine stands of the second age class (21–40 years old) growing in coniferous habitats; and (iv) the equation should include trees with different biosocial positions in line with Kraft’s classification.
2. Materials and Methods
2.1. Description of the Study Site
The research was conducted in single species stands of Scots pine (Pinus sylvestris L.) in the south-western region of Poland. The research area was located between 18°35′ E and 19°20′ E, and 51°00′ N and 51°13′ N. The mean total of precipitation amounted to 641 mm·year−1, with the heaviest precipitation recorded in July (118.5 mm) and the lowest in February (31.7 mm). The average annual temperature was 8.1 °C. July was the warmest (19.2 °C) and January was the coolest (−1.6 °C) month. The growing season lasted for ~222 days [82]. In the managed forests, 20 experimental plots of 0.25 ha (50 × 50 m) were designated. The age of the pine stands ranged from 30 to 38 years (Table 1). In the examined pine stands, there was a lot of dead wood due to density because no intermediate felling was made in them. The stands grew on podzols and brunic arenosols in the habitat conditions typical for pine, i.e., fresh or mixed fresh forest.
2.2. Biometric Measurements
On the designated plots, the diameter at breast height (DBH =) was measured for all 14,935 trees. The DBH of the tree stands was in the range of 9 to 30 cm. This parameter was measured with a metal caliper Mantax Blue (Haglöf, Långsele, Västernorrland, Sweden) in two directions, N-S and W-E, with an accuracy of 0.1 cm, and the arithmetic mean of these measurements was assumed as the DBH. Based on the DBH measurement of all trees on each plot, 15 sample trees were randomly selected, taking into account dominant, codominant, intermediate, and suppressed trees. The selection was made using the English method developed by Humme felling every k-th tree based on the formula [83]:
where: , total number of trees in a research area; , the assumed number of sample trees.
During this felling, sample trees with different DBH, height, and crown length were obtained.
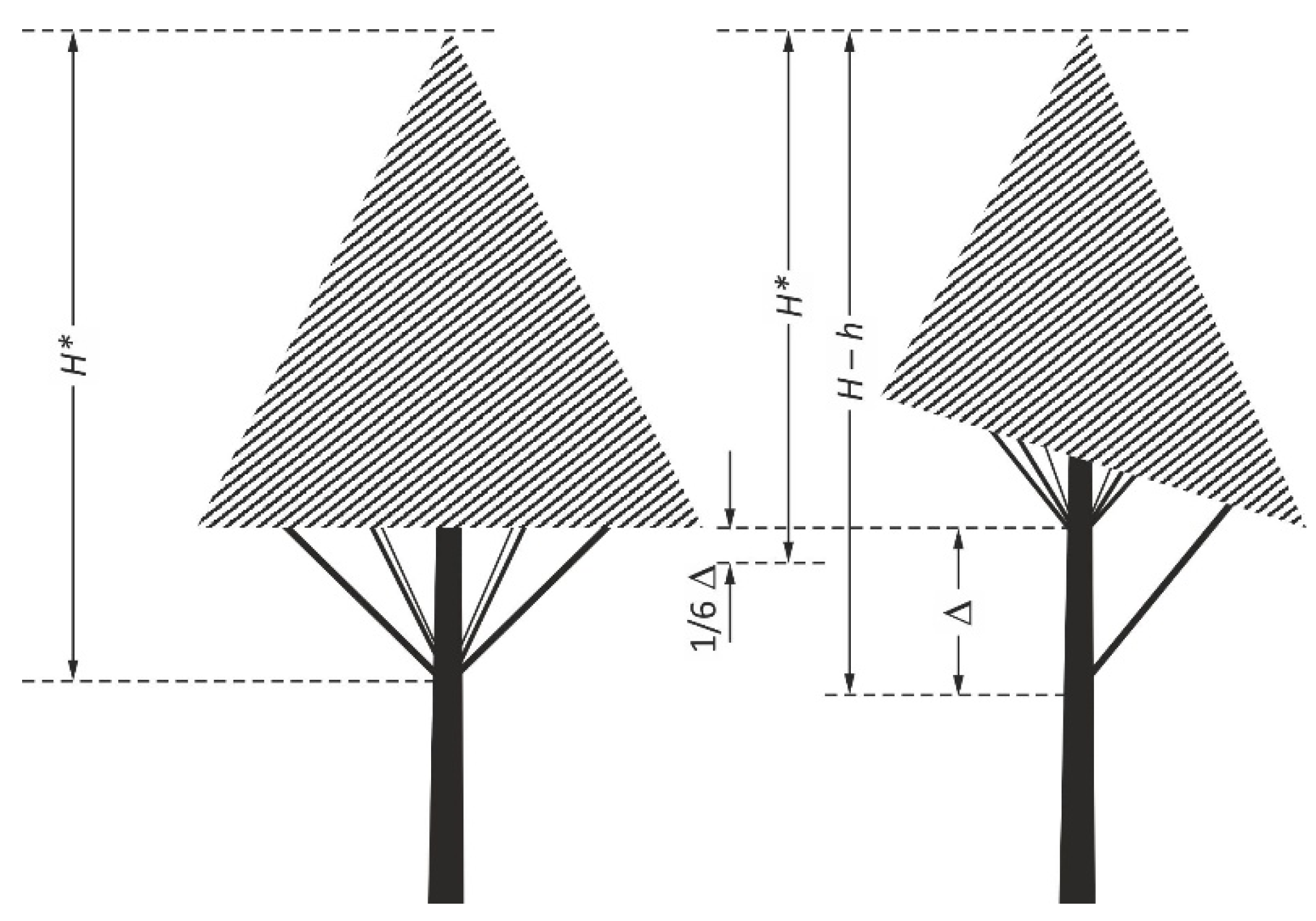
The selected sample trees were felled in March before the start of the growing season. A total of 300 trees were felled. After felling, their length was measured with a metal tape to the nearest 0.01 m and taken as the tree height () and crown length (). In this study, the following method was used to measure the crown length of the trees. A correctly formed whorl of a Scots pine has six branches. In this case, the crown is regular (Figure 1), and its length is defined as the distance from the treetop to the base of the whorl. If in the lowest whorl, one branch (of the size corresponding to the other branches) is dead, the length of the crown is the distance from the treetop to the base of the “incomplete” whorl, minus 1/6 of the distance between this base and the base of the nearest whole whorl (Figure 1). From this assumption, it follows that the crown length is reduced by 1/6 of the annual growth of the stem. In other words, if there are 2 dead branches in the last incomplete whorl, the measurement is shortened by 2/6 of the annual growth.
The above relationship is described by equation [35]:
where: , crown length; H − h, distance from the treetop to the base of the whorl with withered (missing) branches; n, the number of withered (missing) branches in the whorl.
For each plot, the measured or calculated statistics for the stand were as follows: , diameter at breast height (1.3 m above ground level); , trees’ height; , empirical crown length , stand basal area; , the age of trees; , the number of trees per hectare; , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])). The biometric description of the sites is presented in Table 1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of surveyed Scots pine stands.
| Variable | Mean | Minimum | Maximum | Median | Standard Deviation |
|---|---|---|---|---|---|
| (m) | 13.9 | 7.0 | 21.1 | 13.5 | 3.20 |
| (m) | 5.1 | 2.4 | 9.4 | 4.9 | 1.24 |
| (cm) | 13.0 | 6.0 | 24.0 | 13.0 | 3.37 |
| (cm) | 14.7 | 7.7 | 25.7 | 14.7 | 3.37 |
| (trees ha−1) | 2987 | 492 | 8340 | 3174 | 1786.4 |
| (m2 ha−1) | 24.0 | 11.7 | 32.6 | 23.9 | 4.85 |
| (-) | 1.07 | 0.31 | 1.86 | 1.14 | 0.40 |
| (year) | 33 | 30 | 38 | 33 | 2.39 |
, trees height; , empirical crown length; , diameter at breast height (1.3 m above ground level); , the diameter of the trees at the ground level (); , the number of trees per hectare; , stand basal area; , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])); , age of trees.
2.3. Dependency Detection
We assume that there is an allometric relationship between the size of one organ of an individual ( and the size of another part of their body (:
Thus, the allometric relationship between the length of the tree crown and some other measure of its size can be expressed as follows:
We do not know the value of the factor of proportionality , the exponent , or exactly what features fit into the measure . Previous studies [8,60,69,74] have indicated that the measure consists of the thickness of the tree and its height, as well as the stand density [11,85]. Therefore, we are left with a task to develop an allometric model that would allow us to calculate the length of the crown based on the collected empirical data.
3. Results
3.1. Analysis of Data for Building the Crown Length Model
It was assumed that the length of the tree crown in a single species stand (Pinus sylvestris L.) would depend mainly on stand density. The relationships between DBH, height, density, and crown length for 300 sample trees are presented in Table 2.
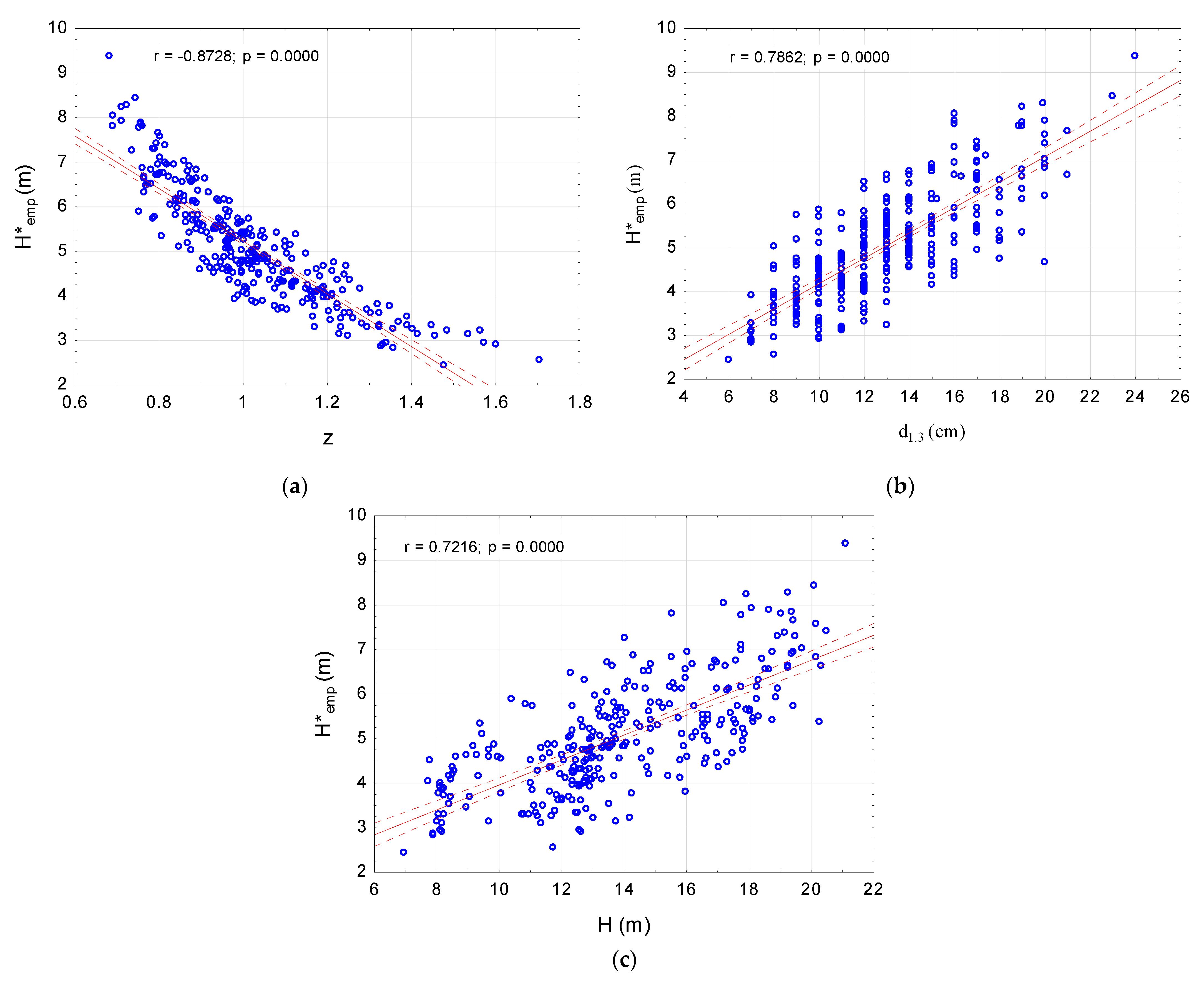
The correlation coefficients ranged from −0.375 to −0.873, and they were clearly higher than the critical values for the assumed significance level of 0.05. The highest correlation coefficient was found for and (r = −0.873; p < 0.0001). In this case, a linear negative correlation was determined. The higher the density of the stand, the shorter the crowns were (Figure 2a). In the case of a positive correlation between and d1.3, the coefficient was r = 0.786; p < 0.0001 (Figure 2b). A slightly weaker relationship was found between and (r = 0.722; p < 0.0001) (Figure 2c).
The density of the studied pine stands differed from 492 to 8340 trees per hectare (Table 3). Biometric parameters of the studied stands ( calculated for 15 sample trees for each experimental plot are summarised in Table 3. Empirical data showed that at the density index of = 1.83, the average crown length was the shortest and equalled = 3.98 m, while at the lowest density of = 0.31, the crown was the longest, amounting to = 6.99 m. The greater the space between the trees, the longer the crown was. For the final dataset, the summary statistics were used to fit the crown length equation (Table 3).
3.2. Crown Length Modelling
Three predictive variables, i.e., DBH, height, and density index (, were used to build the crown length model. The density index was defined as the ratio of the actual number of trees per hectare (which we know only at the time of measurement) to the expected number of trees in a given age and stand quality class (value read from Tables of Stands Volume and Increment [84]). It seems that the introduction of this predictor into the model was essential, as the collected empirical material exhibited a large variation in the number of trees on individual experimental plots.
The analysis of the data in Table 3 confirmed the emerging assumption about the size of measure x in Equation (4):
where: , trees height (m); , DBH (cm); , density index.
Since the diameter at breast heights () is a technical value, it can be replaced by the diameter of the tree at ground level () [85]:
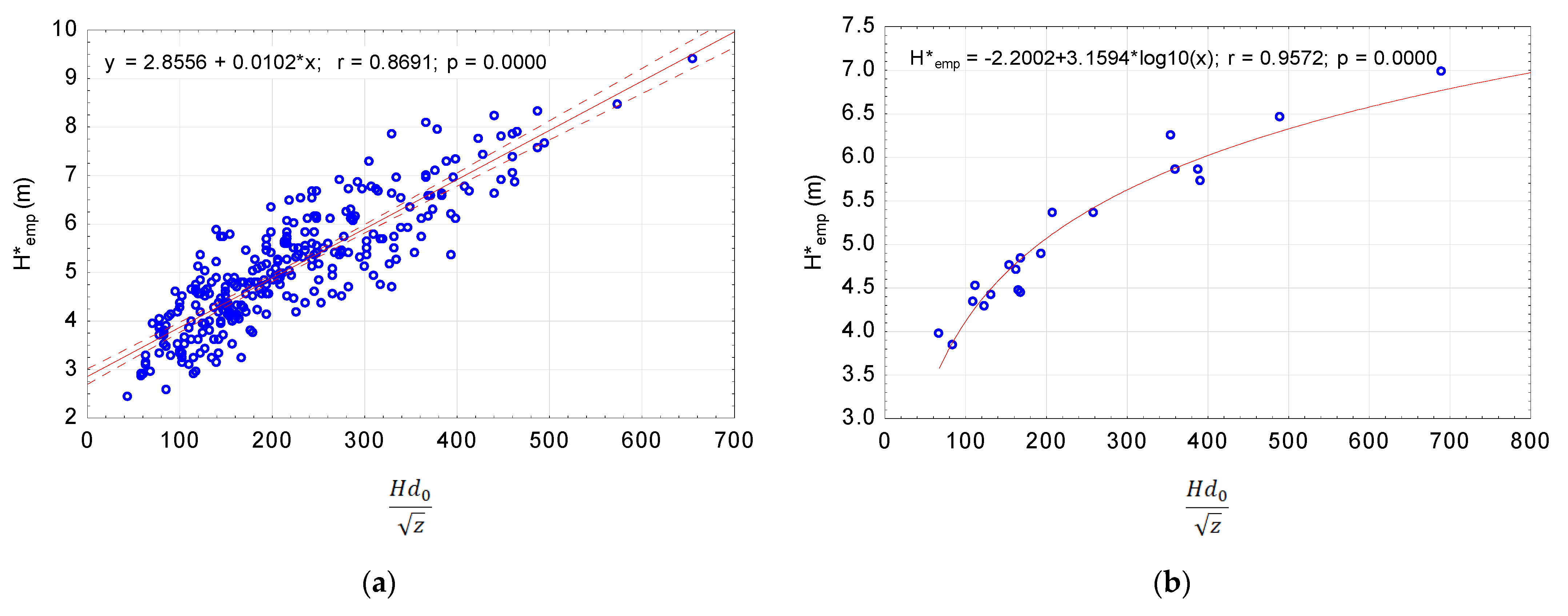
A graphical image of the crown length as a function of density, DBH, and height is illustrated in Figure 3.
The points representing the crown length were arranged around the line according to the equation:
where: , the average calculated crown length (m); , average trees’ height (m); , average diameter of the base of the tree (cm); , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])); , factor of proportionality—dimensionless number (-); .
In the first attempt to calculate the value of the index and the exponent , four stands were selected, i.e., the stand with the highest density index (plot No. 9), maximum height (plot No. 12), maximum DBH (plot No. 11), and the shortest crown (plot No. 17). The parameters of the selected stands are listed in Table 4.
To calculate the values and , the Smolik algorithm was used [86]:
where: ; —are the measured values; , empirical crown lengths (m); is the quotient of the products of the average height and the average diameter of the tree at ground level in the numerator to the root of the density index in the denominator.
The values obtained from the four sample plots, = 0.2505 and = 1.3621, were approximate values. Thus, the equation can be written as follows:
This means that the square of the crown length is directly proportional to the geometric mean height and diameter of the tree at ground level. Such a relationship is logical and seems to reflect the actual situation, where is equivalent to the leaf area index, while the geometric mean height and diameter of the tree at ground level express the effect of the assimilation.
Therefore, adopting , the factor of proportionality was calculated as an average for the twenty studied stands based on the following Equation (12):
This way, the mean value of the factor of proportionality for the stands was obtained (Table 5).
Thus, equation takes the following form:
where: , calculated crowns length (m); H, trees’ height (m); , diameter of the base of the tree (cm); , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])); factor of proportionality .
The value of the factor of proportionality was identical for .
Finally, the model (13) can be represented as:
where: denotes the surface of a cylinder with diameter and height .
In stands, where , i.e., and the density is optimal for the growth and development of trees, Equation (14) has the following form:
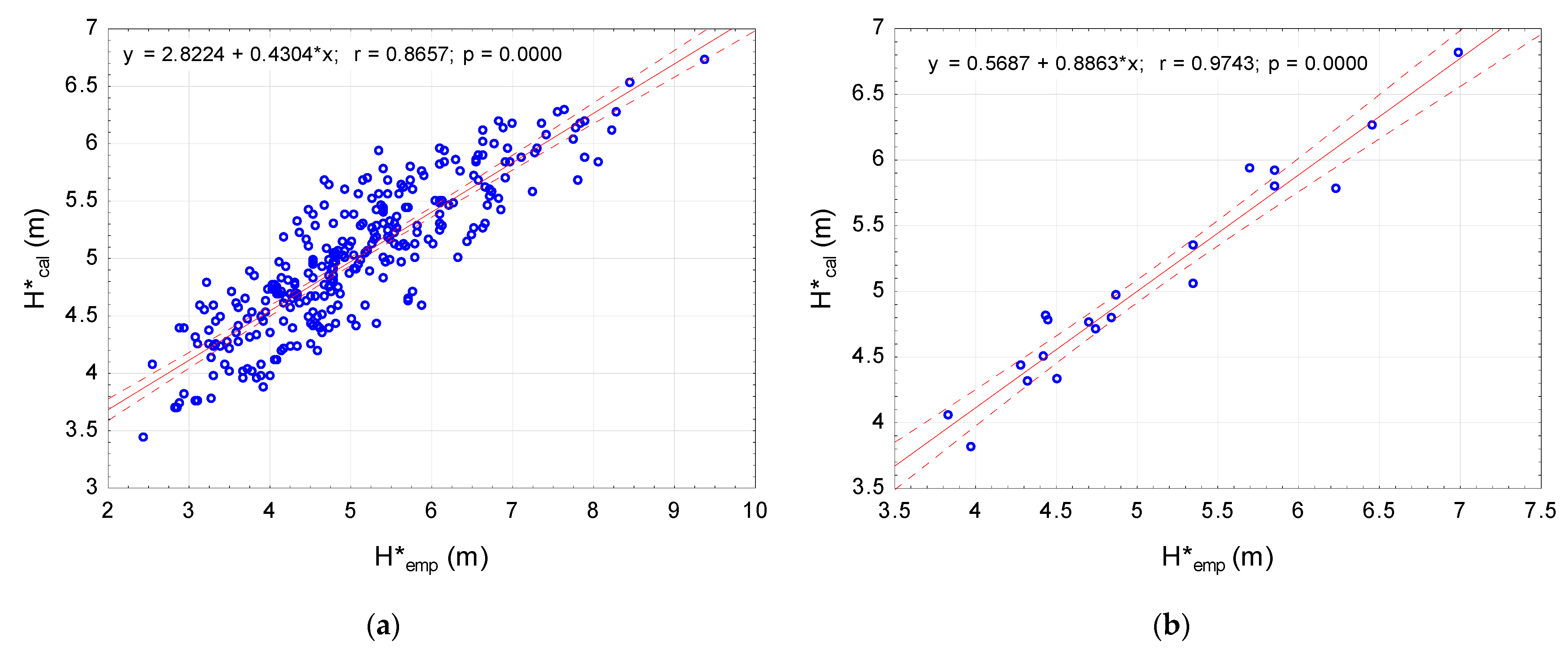
The developed allometric crown length model (13) was used to calculate for the 20 analysed stands. The correlation coefficient between the empirical crown length for the stand () and the calculated model was r = 0.974 (p < 0.0001), with a discrepancy of (±) 3.17% (Table 5). The dispersion of empirical values and the calculated ones for the stands and model trees are presented in Figure 4a,b.
4. Discussion
In many respects, the length of the crown is an interesting biophysical property. However, in forestry surveys, it is rarely measured at both the tree and stand level. This is justified by the fact that the gain from having empirical information is considered lower than the cost of measuring the length of the crown [38]. Therefore, equations that accurately represent actual crown lengths are still being sought. In principle, crown length can be predicted on the basis of standard variables measured in forest management [38,87,88]. However, it is believed that it is very difficult to accurately calculate the length of the crown using basic measurements of trees, i.e., DBH, height, basal area, and stand age, without calibration at the stand level [38]. In our study, the crown length of the sample trees showed a strong correlation with both the DBH (r = 0.786; p < 0.0001) and height (r = 0.722 p < 0.0001) of the trees. Confirmation of such a relationship can be found in the research by Żybura [60], who, on the basis of these features, built tables of stand crown lengths for Pinus sylvestris. The mean error of these tables when determining the absolute crown length was 5.6%. Turski et al. [89] demonstrated that the relationship between the crown length and tree height was weaker than that between the crown length and DBH. However, we must accept the fact that there is much greater variation in crown length between different Pinus sylvestris stands than within a single stand [90]. In our study, the coefficients of variation in the crown length within one stand ranged from 10.8% to 23.3%. However, for all 20 examined stands, the coefficient of variation was higher and amounted to 24.5%. Such a large variation in crown length was influenced by, among others, the density of the examined tree stands. The correlation coefficient between these features was r = −0.873; p < 0.0001 (Table 2). Confirmation of this relationship can be found in the research of other authors [54,78,79,80,81].
Simple crown equations that restrict the predictor variables to one or two measurements of tree size are mostly useful in situations in which the differences in densities from stand to stand are limited while the stand structure is simple [76]. Gill et al. [91] stated that their use of only DBH to estimate the crown radius was likely helped by the fact that their data had been collected primarily from managed forests that had been treated to maintain a low density. Among several tested variables relating to density, studies related to crown allometry identified the basal area as the parameter that best described the effect of density on crown length [71,77].
The proposed crown length model requires the measurement of , , and as input variables. The model shows high compatibility with empirical data within the studied stands (r = 0.974) (Figure 4), which differed in their DBH, height, age, biosocial position, and, above all, density (SD = 1786) (Table 1). The high efficiency of the model indicates the accuracy of its design and the selection of the parameters and These discrepancies come from measurement inaccuracies and incomplete representativeness of the model trees, which could be indicated by a lower coefficient of correlation (r = 0.866; p < 0.0001) for the model trees (Figure 4a). Thus, the model should work better for a community than for a single tree.
It is considered that the most visible changes in crown and trunk size concern data collected from a wide range of stand densities, which could be a problem when fitting equations [76]. It seems, therefore, that the inclusion of the density index as a predictive variable in this model was highly beneficial. The developed model could work well in overpopulated and thinned communities. The analysis showed that “the square of the crown length is directly proportional to the geometric mean height and diameter of the tree at the ground level”. Such a relationship is a representation of biological processes occurring for many years in the phytocenose of pine, which can be expressed as: “the square of the length of the tree crown is equivalent to the leaf area index, and the geometric mean height and diameter express the effect of assimilation”.
The development of the proposed crown length model is based on the assumption that the crown is formed by whorls built of six branches. While measuring the length of the crowns in the sample trees, their length was reduced depending on the number of branches missing from the whorl (Figure 1). This is a new approach to the definition of crown length, as it is based on biological rather than technical considerations. The proposed model allows the crown length to be calculated for both individual trees and stands. DBH, height, and density are the main variables at the tree and stand level, respectively, and are used extensively around the world in forest inventories and forest management planning. This fact speaks for a wider use of the proposed model in forestry practice.
5. Conclusions
In this article, we proposed the allometric model of crown length determination for Pinus sylvestris L., which uses easy-to-measure biometric parameters, i.e., DBH, tree height, and density. It proved highly compatible with the actual crown lengths, as evidenced by the coefficient of determination r2 = 0.95. The model is based on empirical data collected from 300 sample trees growing at different densities (from = 0.31 to = 1.86) and different ages (30 to 38 years). The development of an empirical crown length model is an important task of applied ecology. From an economic point of view, this model can be used to predict the mechanical stability of trees and their susceptibility to wind and snow damage. As it turned out, the equation benefited from including the density index, which mainly determines the amount of light reaching the lower branches of the crown and whether the needles remain on the tree or fall. By developing the allometric crown length model, a very useful indirect tool for estimating foliage size was provided, as it could determine the effectiveness of the physiological processes responsible, among others, for the growth of the tree. The precise determination of the crown length could, therefore, be one of the components used to estimate the mass of needles or leaf area index (LAI) and, consequently, the amount of transpiration or the amount of carbon dioxide bound, which is crucial in the context of climate change. Our further research work will focus on developing models to calculate the mass and leaf area index, as well as the amount of transpiration in pine monocultures, and the crown length model will be one of the most important components in this modelling.
Author Contributions
Conceptualisation, M.S. and K.S.; Methodology, M.S. and K.S.; Formal Analysis, M.S. and K.S.; Investigation, M.S.; Resources, M.S.; Data Curation, M.S.; Writing—Original Draft Preparation, M.S.; Writing—Review and Editing, M.S. and K.S.; Visualisation, M.S.; Supervision, M.S. and K.S.; Project Administration, M.S. and K.S.; Funding Acquisition, M.S. and K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hytteborn, H.; Maslov, A.A.; Nazimova, D.I.; Rysin, L.P. Boreal forests of Eurasia. In Ecosystems of the World 6. Coniferous Forests; Andersson, F., Ed.; Elsevier: Amsterdam, The Netherland, 2005; Volume 6, pp. 23–99. [Google Scholar]
- Labra, M.; Grassi, F.; Sgorbati, S.; Ferrari, C. Distribution of genetic variability in southern populations of Scots pine (Pinus sylvestris L.) from the Alps to the Apennines. Flora Morphol. Distrib. Funct. Ecol. Plants 2006, 201, 468–476. [Google Scholar] [CrossRef]
- Statistical Yearbook of Forestry; Statistics Poland: Warszawa, Poland, 2022. Available online: https://stat.gov.pl (accessed on 3 August 2023).
- Assmann, E. The Principles of Forest Yield Study; Pergamon Press: Oxford, UK, 1970; p. 506. [Google Scholar]
- Smith, D.M. 1986 The Practice of Silviculture, 8th ed.; John Wiley and Sons, Inc.: New York, NY, USA, 1986; p. 527. [Google Scholar]
- Hann, D.W.; Wang, C.H. Mortality Equations for Individual Trees in Southwest Oregon; Forest Research Lab., Oregon State University, Corvallis, Research Bulletin: Corvallis, OR, USA, 1990; Volume 67, pp. 1–17. [Google Scholar]
- Valentine, H.T.; Ludlow, A.R.; Furnival, G.M. Modelling crown rise in even-aged stands of Sitka spruce or loblolly pine. For. Ecol. Manag. 1994, 69, 189–197. [Google Scholar] [CrossRef]
- Hasenauer, H.; Monserud, R.A. A crown ratio model for Austrian forests. For. Ecol. Manag. 1996, 84, 49–60. [Google Scholar] [CrossRef]
- Monserud, R.A.; Sterba, H. A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Jack, S.B.; Long, J.N. Forest production and the organization of foliage within crowns and canopies. Forest Ecol. Manag. 1992, 49, 233–245. [Google Scholar] [CrossRef]
- Sporek, M. The Leaf Area of Scots Pine (Pinus sylvestris L.) as a Function of the Population Structure; PWN: Warszawa, Poland, 2018; p. 208. ISBN 978-83-01-20366-5. (In Polish) [Google Scholar]
- Maguire, D.A.; Kershaw, J.A., Jr.; Hann, D.W. Predicting the effects of silvicultural regime on branch size and crown wood core in Douglas-fir. For. Sci. 1991, 37, 1409–1428. [Google Scholar] [CrossRef]
- Kellomäki, S.; Ikonen, V.; Kolstrom, T. Modelling the structural growth of Scots pine with implications for wood quality. Ecol. Model. 1999, 122, 117–134. [Google Scholar] [CrossRef]
- Rikala, J. Spruce and Pine on Drained Peatlands—Wood Quality and 1 Suitability for the Sawmill Industry; University of Helsinki, Department of Forest Resource Management, Publications 35: Helsinki, Finland, 2003; p. 147. [Google Scholar]
- Valentine, H.T.; Mäkelä, A. Bridging process-based and empirical approaches to modeling tree growth. Tree Physiol. 2005, 25, 769–779. [Google Scholar] [CrossRef]
- Maltamo, M.; Peuhkurinen, J.; Malinen, J.; Vauhkonen, J.; Packalén, P.; Tokola, T. Predicting tree attributes and quality characteristics of Scots pine using airborne laser scanning data. Silva Fenn. 2009, 43, 507–521. Available online: http://www.metla.fi/silvafennica/full/sf43/sf433507.pdf (accessed on 8 August 2023). [CrossRef]
- Wilson, J.S.; Oliver, C.D. Stability and density management in Douglas-fir plantations. Can. J. For. Res. 2000, 30, 910–920. [Google Scholar] [CrossRef]
- Leersnijder, R.P. Pinogram: A Pine Growth Area Model. Ph.D. Thesis, Wageningen University and Research, Department of Forestry, WAU, Wageningen, The Netherlands, 1992; p. 147, ISBN 9054850027. Available online: https://edepot.wur.nl/202715 (accessed on 1 March 2023).
- Gardiner, B.A.; Peltola, H.; Kellomaki, S. Comparison of two models for predicting the critical wind speed required to damage coniferous trees. Ecol. Mod. 2000, 129, 1–23. [Google Scholar] [CrossRef]
- Moore, J. Mechanical Behavior of Coniferous Trees Subjected to Wind Loading. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2002. Available online: Moore_John_R_2002.pdf (accessed on 20 March 2023).
- Cucchi, V.; Meredieu, C.; Stokes, A.; de Coligny, F.; Suarez, J.; Gardiner, B.A. Modelling the windthrow risk for simulated forest stands of maritime pine (Pinus pinaster Ait.). For. Ecol. Manag. 2005, 213, 184–196. [Google Scholar] [CrossRef]
- van Wagner, C.E. Conditions for the start and spread of crown fire. Can. J. For. Res. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Keyes, C.R.; O’Hara, K.L. Quantifying stand targets for silvicultural prevention of crown fires. West. J. Appl. For. 2002, 17, 101–109. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Rigolot, E. The fire ecology and management of maritime pine (Pinus pinaster Ait). For. Ecol. Manag. 2007, 241, 1–13. [Google Scholar] [CrossRef]
- Sporek, M.; Sporek, K.; Stebila, J.; Kučerka, M.; Kminiak, R.; Lubis, M.A.R. Assessment of the mass and surface area of the Scots pine (Pinus sylvestris L.) needles. Appl. Sci. 2022, 12, 8204. [Google Scholar] [CrossRef]
- Burger, H. Wood, number of leaves and growth. I. The Weymouth pine. Mitt. Schweiz. Anst. Forstl. Versuchsw. 1929, 15, 243–292. (In German) [Google Scholar]
- Burger, H. Crown structure of coniferous stands of the same age. Mitt. Schweiz. Anst. Forstl. Versuch. 1939, 21, 5–58. (In German) [Google Scholar]
- Burger, H. Tree crown and growth in two mature stands of spruce. Mitt. Schweiz. Anst. Forstl. Versuchsw. 1939, 21, 147–176. (In German) [Google Scholar]
- Burger, H. 1941. Wood, number of leaves and growth. V. Spruces and pines of different origins at different sites. Mitt. Schweiz. Anst. Forstl. Versuchsw. 1941, 22, 10–60. (In German) [Google Scholar]
- Burger, H. Wood, number of leaves and growth. IX. The pine. Mitt. Schweiz. Anst. Forstl. Versuchsw. 1947, 25, 435–493. (In German) [Google Scholar]
- Burger, H. Wood, number of leaves and growth. XIII. Spruces in the tall forest of the same age. Mitt. Schweiz. Anst. Forstl. Versuch. 1953, 29, 38–130. (In German) [Google Scholar]
- Lemke, J. The crown as a criterion for assessing the growth dynamics of trees in a pine stand. Fol. For. Pol. 1966, 12, 185–236. (In Polish) [Google Scholar]
- Lemke, J. Relationship between the size of the crown and the increment of trees in pine stands. Pr. Kom. Nauk Roln. Kom. Nauk Leśn. PTPN 1968, 25, 1–48. (In Polish) [Google Scholar]
- Lemke, J. The usefulness of the crown and height of trees for estimating their incremental properties in pine stands of the 2nd and 3rd age class. Pr. Kom. Nauk Roln. Kom. Nauk Leśn. PTPN 1971, 32, 73–87. (In Polish) [Google Scholar]
- Sporek, K. Length of Trees-Crown as Function of Structure of Scots Pine Even-Aged Forest Stands; PWN Warszawa: Wrocław, Poland, 1990; p. 31. ISBN 83-01-10228-4. (In Polish) [Google Scholar]
- Ducey, M.J. Predicting crown size and shape from simple stand variables. J. Sust. For. 2009, 28, 5–21. [Google Scholar] [CrossRef]
- Urban, J.; Tatarinov, F.; Nadezhdina, N.; Cermak, J.; Ceulemans, R. Crown structure and leaf area of the understorey species Prunus serotina. Trees—Struct. Funct 2009, 23, 391–399. [Google Scholar] [CrossRef]
- Maltamo, M.; Bollandsås, O.M.; Vauhkonen, J.; Breidenbach, J.; Gobakken, T.; Næsset, E. Comparing different methods for prediction of mean crown height in Norway spruce stands using airborne laser scanner data. Forestry 2010, 83, 257–268. [Google Scholar] [CrossRef]
- Gering, L.R.; May, D.M. The relationship of diameter at breast height and crown diameter for four species in Hardin County, Tennessee. South. J. Appl. For. 1995, 19, 177–181. [Google Scholar] [CrossRef]
- Foli, E.G.; Alder, D.; Miller, H.G.; Swaine, M.D. Modelling growing space requirements for some tropical forest tree species. For. Ecol. Manag. 2003, 173, 79–88. [Google Scholar] [CrossRef]
- Condes, S.; Sterba, H. Derivation of compatible crown width equations for some important tree species of Spain. For. Ecol. Manag. 2005, 217, 203–218. [Google Scholar] [CrossRef]
- Dubravac, T. Application of digitalisation of crowns and method of visualisation in a study of stand structures. Rad. Sumar. 2005, 40, 53–72. (In Croatian) [Google Scholar]
- Hemery, G.E.; Savill, P.S.; Pryors, S.N. Application of the crown diameter−stem diameter relationship for different species of broadleaved trees. For. Ecol. Manag. 2005, 215, 285–294. [Google Scholar] [CrossRef]
- Lockhart, B.R.; Robert, C.; Weih, J.R.; Keith, M.S. Crown radius and diameter at breast height relationships for six bottomland hardwood species. J. Ark. Acad. Sci. 2005, 59, 110–115. [Google Scholar]
- Sánchez-González, M.; Cañellas, I.; Montero, G. Generalized height-diameter and crown diameter prediction models for cork oak forests in Spain. For. Syst. 2007, 16, 76–88. [Google Scholar] [CrossRef]
- Dubravac, T.; Dekanić, S.; Vrbek, B.; Matošević, D.; Roth, V.; Jakovlević, T.; Zlatanov, T. Crown volume in forest stands of pedunculate oak and common hornbeam. Period. Biol. 2009, 111, 479–485. Available online: https://hrcak.srce.hr/file/73942 (accessed on 1 March 2023).
- Russell, M.B.; Weiskittel, A.R. Maximum and Largest Crown Width Equations for 15 Tree Species in Maine. North Jersey App. For. 2011, 28, 84–91. [Google Scholar] [CrossRef]
- Fu, L.; Sun, H.; Sharma, R.P.; Lei, Y.; Zhang, H.; Tang, S. Nonlinear mixed-effects crown width models for individual trees of Chinese fir (Cunninghamia lanceolata) in south-central China. For. Ecol. Manag. 2013, 302, 210–220. [Google Scholar] [CrossRef]
- Shama, R.P.; Vacek, A.; Vacek, S. Individual tree crown width models for Norway spruce and European beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]
- Sharma, R.P.; Bílek, L.; Vacek, Z.; Vacek, S. Modelling crown width-diameter relationship for Scots pine in the central Europe. Trees 2017, 31, 1875–1889. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, S. Allometric modelling of crown width for white spruce by fixed- and mixed-effects models. For. Chron. 2017, 93, 138–147. [Google Scholar] [CrossRef]
- Raptis, D.; Kazana, V.; Kazaklis, A.; Stamatiou, C.A. Crown Width-Diameter Model for Natural Even-Aged Black Pine Forest Management. Forests 2018, 9, 610. [Google Scholar] [CrossRef]
- Qiu, S.; Liang, R.; Wang, Y.; Luo, M.; Sun, Y. Comparative analysis of machine learning algorithms and statistical models for predicting crown width of Larix olgensis. Earth Sci. Inform. 2022, 15, 2415–2429. [Google Scholar] [CrossRef]
- Qiu, S.; Gao, P.; Pan, L.; Zhou, L.; Liang, L.; Sun, Y.; Wang, Y. Developing nonlinear additive tree crown width models based on decomposed competition index and tree variables. J. For. Res. 2023, 34, 1407–1422. [Google Scholar] [CrossRef]
- Bella, I.E. A new competition model for individual trees. For. Sci. 1971, 17, 364–372. [Google Scholar] [CrossRef]
- Clutter, J.L.; Fotson, J.C.; Pienaar, L.V.; Brister, G.H.; Bailey, R.L. Timber Management: A Quantitative Approach; John Wiley and Sons: Now York, NY, USA, 1983; p. 333. [Google Scholar]
- Sprinz, P.T.; Burkhart, H.E. Relationships between tree crown, stem and stand characteristics in unthinned loblolly pine plantations. Can. J. For. Res. 1987, 17, 534–538. [Google Scholar] [CrossRef]
- Temesgen, H.; LeMay, V.; Mitchell, S.J. Tree crown ratio models for multi-species and multi-layered stands of southeastern British Columbia. For. Chron. 2005, 81, 133–141. [Google Scholar] [CrossRef]
- Borowski, M. Przyrost Drzew i Drzewostanów; PWRiL: Warszawa, Poland, 1974. (In Polish) [Google Scholar]
- Żybura, H. Lenght of tree crowns in pine stands. Sylwan 1977, 1, 13–20. (In Polish) [Google Scholar]
- Grochowski, J. Accuracy of volume measurement according to Denzin and the biological structure of the stand expressed by distribution curves of the relative length of the crown. Pr. I Spraw. Zakładu Urządzenia Lasu SGGW W Warszawie 1934, 81, 343–386. (In Polish) [Google Scholar]
- Hatch, C.R.; Gerrard, D.J.; Tappeiner, J.C., II. Exposed crown surface area: A mathematical index of individual tree growth potential. Can. J. For. Res. 1975, 5, 224–228. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grote, R.; Reineking, B.; Rötzer, T.; Seifert, S. Models for forest ecosystem management: A European perspective. Ann. Bot. 2008, 101, 1065–1087. [Google Scholar] [CrossRef]
- Kändler, G. Die Determination of Canopy Parameters as Input Variables for Interception Models Using Aerophotogrammetric Methods; Mitteilungen der Forstlichen Versuchs- und Forschungsanstalt: Freiburg, Germany, 1986; p. 129. (In German) [Google Scholar]
- Mohren, G.M.J. Simulation of Forest Growth, Applied to Douglas-fir Stands in the Netherlands. Ph.D. Thesis, Agricultural University of Wagenin, Wagenin, The Netherlands, 1987; p. 201257. Available online: wur.nl (accessed on 20 July 2022).
- Biging, G.S.; Wensel, L.C. Estimation of crown form for six conifer species of northern California. Can. J. For. Res. 1990, 20, 1137–1142. [Google Scholar] [CrossRef]
- Hann, D.W. An adjustable predictor of crown profile for stand-grown Douglas-fir trees. For. Sci. 1999, 45, 217–225. [Google Scholar] [CrossRef]
- Rautiainen, M.; Stenberg, P. Simplified tree crown model using standard forest mensuration data for Scots pine. Agric. For. Meteorol. 2005, 128, 123–129. [Google Scholar] [CrossRef]
- Baldwin, V.C., Jr.; Peterson, K.D. Predicting the crown shape of loblolly pine trees. Can. J. For. Res. 1997, 27, 102–107. [Google Scholar] [CrossRef]
- Doruska, P.F.; Mays, J.E. Crown profile modeling of loblolly pine by nonparametric regression analysis. For. Sci. 1998, 44, 445–453. [Google Scholar] [CrossRef]
- Holdaway, M.R. Modeling tree crown ratio. For. Chron. 1986, 62, 451–455. [Google Scholar] [CrossRef]
- Dyer, M.E.; Burkhart, H.E. Compatible crown ratio and crown height models. Can. J. For. Res. 1987, 17, 572–574. [Google Scholar] [CrossRef]
- Zhang, S.; Burkhart, H.; Amateis, R. Modeling individual tree growth for juvenile loblolly pine plantations. For. Ecol. Manag. 1996, 89, 157–172. [Google Scholar] [CrossRef]
- Gilmore, D.W. Equations to describe crown allometry of Larix require local validation. For. Ecol. Manag. 2001, 148, 109–116. [Google Scholar] [CrossRef]
- Crecente-Campo, F. Single Tree Growth Model for Pinus Radiata D. Don in Galicia. Ph.D. Thesis, University of Santiago de Compostela, Lugo, Spain, 2008. Available online: http://hdl.handle.net/10347/2415 (accessed on 15 July 2023).
- Sattler, D.F.; LeMay, V. A system of nonlinear simultaneous equations for crown length and crown radius for the forest dynamics model SORTIE-ND. Can. J. For. Res. 2011, 41, 1567–1576. [Google Scholar] [CrossRef]
- Hynynen, J. Predicting tree crown ratio for unthinned and thinned Scots pine stands. Can. J. For. Res. 1995, 25, 57–62. [Google Scholar] [CrossRef]
- Astrup, R. Modelling Growth of Understory Aspen and Spruce in Western Boreal Canada. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2006. [Google Scholar]
- Rouvinen, S.; Kuuluvainen, T. Structure and asymmetry of tree crowns in relation to local competition in a natural mature Scots pine forest. Can. J. For. Res. 1997, 27, 890–902. [Google Scholar] [CrossRef]
- Meng, S.X.; Lieffers, V.J.; Huang, S.M. Modelling crown volume of lodgepole pine based upon the uniform stress theory. For. Ecol. Manag. 2007, 251, 174–181. [Google Scholar] [CrossRef]
- Seidel, D.; Leuschner, C.; Muller, A.; Krause, B. Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning. For. Ecol. Manag. 2011, 261, 2123–2132. [Google Scholar] [CrossRef]
- Institute of Meteorology and Water Management, National Research Institute. Available online: https://danepubliczne.imgw.pl/data/dane_pomiarowo_obserwacyjne/dane_meteorologiczne/miesieczne/ (accessed on 1 August 2023).
- Czarnowski, M.S. Investigation of the potential productive capacity of forest sites as a function of soil, climate and ecological properties of tree species. (A proposal of standardized method for the world-wide gathering action). Acta Univ. Wratisl. Pr. Bot. 1971, 134, 127–151. [Google Scholar]
- Szymkiewicz, B. Volume and Increment Stands Tables; PWRiL: Warszawa, Poland, 1961. (In Polish) [Google Scholar]
- Czarnowski, M.S. Outline of Terrestrial Plant Ecology; PWN: Warsaw, Poland, 1989; p. 555. (In Polish) [Google Scholar]
- Smolik, S. Equalizing Calculus for Non-Linear Functions; Centrum Obliczeniowe PAN: Warszawa, Poland, 1973. (In Polish) [Google Scholar]
- Ojansuu, R.; Maltamo, M. Sapwood and heartwood taper in Scots pine stems. Can. J. For. Res. 1995, 25, 1928–1943. [Google Scholar] [CrossRef]
- Uusitalo, J.; Kivinen, V.P. Constructing bivariate dbh/dead-branch distribution of pines for use in sawing production planning. Scand. J. For. Res. 1998, 3, 509–514. [Google Scholar] [CrossRef]
- Turski, M.; Jaszczak, R.; Deus, R. Selected features of tree crowns and their relationship with the dbh and height in pine tree−stands of different age classes. Sylwan 2012, 156, 369–378. (In Polish) [Google Scholar]
- Mäkinen, H.; Maltamo, M. Models describing tree quality. In Metsää Kuvaavat Mallit; Maltamo, M., Laukkanen, S., Eds.; Silva Carelica, University of Joensuu, Faculty of Forestry: Joensuu, Finland, 2001; Volume 36, pp. 111–133. (In Finnish) [Google Scholar]
- Gill, S.J.; Biging, G.S.; Murphy, E.C. Modeling conifer tree crown radius and estimating canopy cover. For. Ecol. Manag. 2000, 126, 405–416. [Google Scholar] [CrossRef]
Figure 1.
Clarification of the concept of pine crown length [35].
Figure 1.
Clarification of the concept of pine crown length [35].

Figure 2.
The relationship between: (a) Density index and crown length (b) Diameter at breast height and crown length; (c) Height and crown length. Biometrics for 300 sample trees.
Figure 2.
The relationship between: (a) Density index and crown length (b) Diameter at breast height and crown length; (c) Height and crown length. Biometrics for 300 sample trees.

Figure 3.
Pine crown length as a function of density, diameter at breast height, and height: (a) A scatter plot for 300 model trees; (b) A scatter plot with a matching logarithmic function for 20 stands.
Figure 3.
Pine crown length as a function of density, diameter at breast height, and height: (a) A scatter plot for 300 model trees; (b) A scatter plot with a matching logarithmic function for 20 stands.

Figure 4.
The relationship between: (a) Empirical crown length () and calculated () (for 300 Pinus sylvestris L. model trees); (b) Empirical crown length () and calculated () (for 20 stands of Pinus sylvestris L.).
Figure 4.
The relationship between: (a) Empirical crown length () and calculated () (for 300 Pinus sylvestris L. model trees); (b) Empirical crown length () and calculated () (for 20 stands of Pinus sylvestris L.).

Table 2.
Pearson correlation matrix between crown length and biometric features and density of 300 sample trees. Statistically significant results for p < 0.05.
Table 2.
Pearson correlation matrix between crown length and biometric features and density of 300 sample trees. Statistically significant results for p < 0.05.
| z | |||||
|---|---|---|---|---|---|
| (m) | (cm) | (m) | (-) | ||
| (cm) | Pearson’s r | 0.7862 | – | ||
| p-value | <0.0001 | – | |||
| (m) | Pearson’s r | 0.7216 | 0.8536 | – | |
| p-value | <0.0001 | <0.001 | – | ||
| (-) | Pearson’s r | −0.8728 | −0.4735 | −0.3751 | – |
| p-value | <0.0001 | <0.0001 | <0.0001 | – |
, empirical crowns length; , diameter at breast height (1.3 m above ground level); , trees height; , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])).
Table 3.
Summary statistics of the final dataset used to fit the crown length equation (n = 300 trees measured in 20 plots).
Table 3.
Summary statistics of the final dataset used to fit the crown length equation (n = 300 trees measured in 20 plots).
| Plot Number | ||||||
|---|---|---|---|---|---|---|
| (cm) | (m) | (m) | (-) | (Trees/ha) | (Year) | |
| 1 | 10.5 | 9.70 | 4.51 | 1.12 | 4180 | 30 |
| 2 | 12.2 | 13.41 | 4.84 | 1.23 | 3208 | 30 |
| 3 | 12.3 | 12.64 | 4.46 | 1.15 | 3192 | 31 |
| 4 | 9.3 | 12.52 | 4.29 | 1.25 | 3300 | 33 |
| 5 | 10,9 | 12.34 | 4.42 | 1.40 | 3628 | 34 |
| 6 | 12.5 | 13.41 | 4.88 | 0.97 | 2312 | 35 |
| 7 | 12.5 | 13.43 | 4.71 | 1.36 | 3544 | 32 |
| 8 | 12.7 | 13.95 | 5.35 | 0.93 | 2512 | 30 |
| 9 | 10.7 | 12.06 | 4.33 | 1.86 | 4928 | 35 |
| 10 | 15.6 | 16.97 | 5.35 | 1.27 | 2268 | 38 |
| 11 | 18.5 | 18.96 | 6.46 | 0.61 | 860 | 38 |
| 12 | 18.3 | 19.18 | 6.99 | 0.31 | 492 | 33 |
| 13 | 14.5 | 16.38 | 5.86 | 0.54 | 1188 | 33 |
| 14 | 16.0 | 17.60 | 5.71 | 0.63 | 1252 | 32 |
| 15 | 15.7 | 15.50 | 5.86 | 0.48 | 1092 | 33 |
| 16 | 8.7 | 8.36 | 3.84 | 1.04 | 5080 | 30 |
| 17 | 8.9 | 8.61 | 3.98 | 1.83 | 8340 | 31 |
| 18 | 16.1 | 18.27 | 6.24 | 0.84 | 1468 | 35 |
| 19 | 13.1 | 13.12 | 4.44 | 1.30 | 3156 | 35 |
| 20 | 11.7 | 12.94 | 4.75 | 1.23 | 3740 | 32 |
| Mean | 13.0 | 13.97 | 5.06 | 1.7 | 2987 | - |
, average diameter at breast height (1.3 m above ground level); , average trees height; , average empirical crowns length; , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])); , age of trees; , the number of trees per hectare.
Table 4.
Biometric parameters of four selected stands for determining the factor of proportionality and exponent using the Smolik algorithm [86].
Table 4.
Biometric parameters of four selected stands for determining the factor of proportionality and exponent using the Smolik algorithm [86].
| Plot Number | ||||
|---|---|---|---|---|
| (m) | (cm) | (-) | (m) | |
| 17 | 8.61 | 10.06 | 1.83 | 3.98 |
| 9 | 12.06 | 12.40 | 1.86 | 4.33 |
| 11 | 18.96 | 20.20 | 0.61 | 6.46 |
| 12 | 19.18 | 20.00 | 0.31 | 6.99 |
, average trees height; , average diameter of the base of the tree, , average empirical crowns length; , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])).
Table 5.
Calculated value of the factor of proportionality () for all empirical material and determination of the efficiency of the developed allometric crown length model ().
Table 5.
Calculated value of the factor of proportionality () for all empirical material and determination of the efficiency of the developed allometric crown length model ().
| Plot Number | Divergence | ||||||
|---|---|---|---|---|---|---|---|
| (-) | (cm) | (m) | (m) | (-) | (m) | (%) | |
| 1 | 1.12 | 12.2 | 9.70 | 4.51 | 1.39 | 4.3 | 4.10 |
| 2 | 1.23 | 13.9 | 13.41 | 4.84 | 1.34 | 4.8 | 1.06 |
| 3 | 1.15 | 14.0 | 12.64 | 4.46 | 1.24 | 4.8 | 6.88 |
| 4 | 1.25 | 11.0 | 12.52 | 4.29 | 1.29 | 4.4 | 3.28 |
| 5 | 1.40 | 12.6 | 12.34 | 4.42 | 1.31 | 4.5 | 1.88 |
| 6 | 0.97 | 14.2 | 13.41 | 4.88 | 1.31 | 5.0 | 1.63 |
| 7 | 1.36 | 14.2 | 13.43 | 4.71 | 1.32 | 4.8 | 0.98 |
| 8 | 0.93 | 14.4 | 13.95 | 5.35 | 1.41 | 5.1 | 5.56 |
| 9 | 1.86 | 12.4 | 12.06 | 4.33 | 1.34 | 4.3 | 0.60 |
| 10 | 1.27 | 17.3 | 16.97 | 5.35 | 1.33 | 5.3 | 0.13 |
| 11 | 0.61 | 20.2 | 18.96 | 6.46 | 1.37 | 6.3 | 3.12 |
| 12 | 0.31 | 20.0 | 19.18 | 6.99 | 1.36 | 6.8 | 2.52 |
| 13 | 0.54 | 16.2 | 16.38 | 5.86 | 1.34 | 5.8 | 1.06 |
| 14 | 0.63 | 17.7 | 17.60 | 5.71 | 1.28 | 5.9 | 3.67 |
| 15 | 0.48 | 17.4 | 15.50 | 5.86 | 1.32 | 5.9 | 0.81 |
| 16 | 1.04 | 10.4 | 8.36 | 3.84 | 1.26 | 4.0 | 5.24 |
| 17 | 1.83 | 10.6 | 8.61 | 3.98 | 1.39 | 3.8 | 4.23 |
| 18 | 0.84 | 17.8 | 18.27 | 6.24 | 1.44 | 5.8 | 7.49 |
| 19 | 1.30 | 14.8 | 13.12 | 4.44 | 1.23 | 4.8 | 8.21 |
| 20 | 1.23 | 12.2 | 12.94 | 4.75 | 1.34 | 4.7 | 0.99 |
| Mean | 1.7 | 14.7 | 13.97 | 5.06 | 1.33 | 5.1 | 3.17 |
, age of trees; , density index (the actual number of trees per ha/normative number of trees (taken from Tables of Stands Volume and Increment [84])); , average diameter of the trees at ground level; , average trees height; , average empirical crowns length; , average calculated crowns length; , factor of proportionality.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sporek, M.; Sporek, K. Allometric Model of Crown Length for Pinus sylvestris L. Stands in South-Western Poland. Forests 2023, 14, 1779. https://doi.org/10.3390/f14091779
AMA Style
Sporek M, Sporek K. Allometric Model of Crown Length for Pinus sylvestris L. Stands in South-Western Poland. Forests. 2023; 14(9):1779. https://doi.org/10.3390/f14091779
Chicago/Turabian StyleSporek, Monika, and Kazimierz Sporek. 2023. "Allometric Model of Crown Length for Pinus sylvestris L. Stands in South-Western Poland" Forests 14, no. 9: 1779. https://doi.org/10.3390/f14091779
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.