
Growth Response of Norway Spruce (Picea abies [L.] Karst.) in Central Bohemia (Czech Republic) to Climate Change
 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | GPS (Coordinates) | Altitude (m) | Exposure | Slope (%) | Age (Years) | Height (m) | DBH (cm) | Forest (Type *) |
|---|---|---|---|---|---|---|---|---|
| Karlstejn 1 | 49°56′51.3′′ N, 14°12′05.6′′ E | 422 | N-W | 5 | 88 | 25 | 34 | 3B |
| Cukrak 2 | 49°56′14.2′′ N, 14°21′13.4′′ E | 402 | N-W | 5 | 82 | 25 | 30 | 3K |
| Kostelec 3 | 49°57′55.9′′ N, 14°48′58.9′′ E | 423 | N-W | 5 | 113 | 31 | 41 | 3S |
2.2. Data Collection
2.3. Data Analysis
3. Results
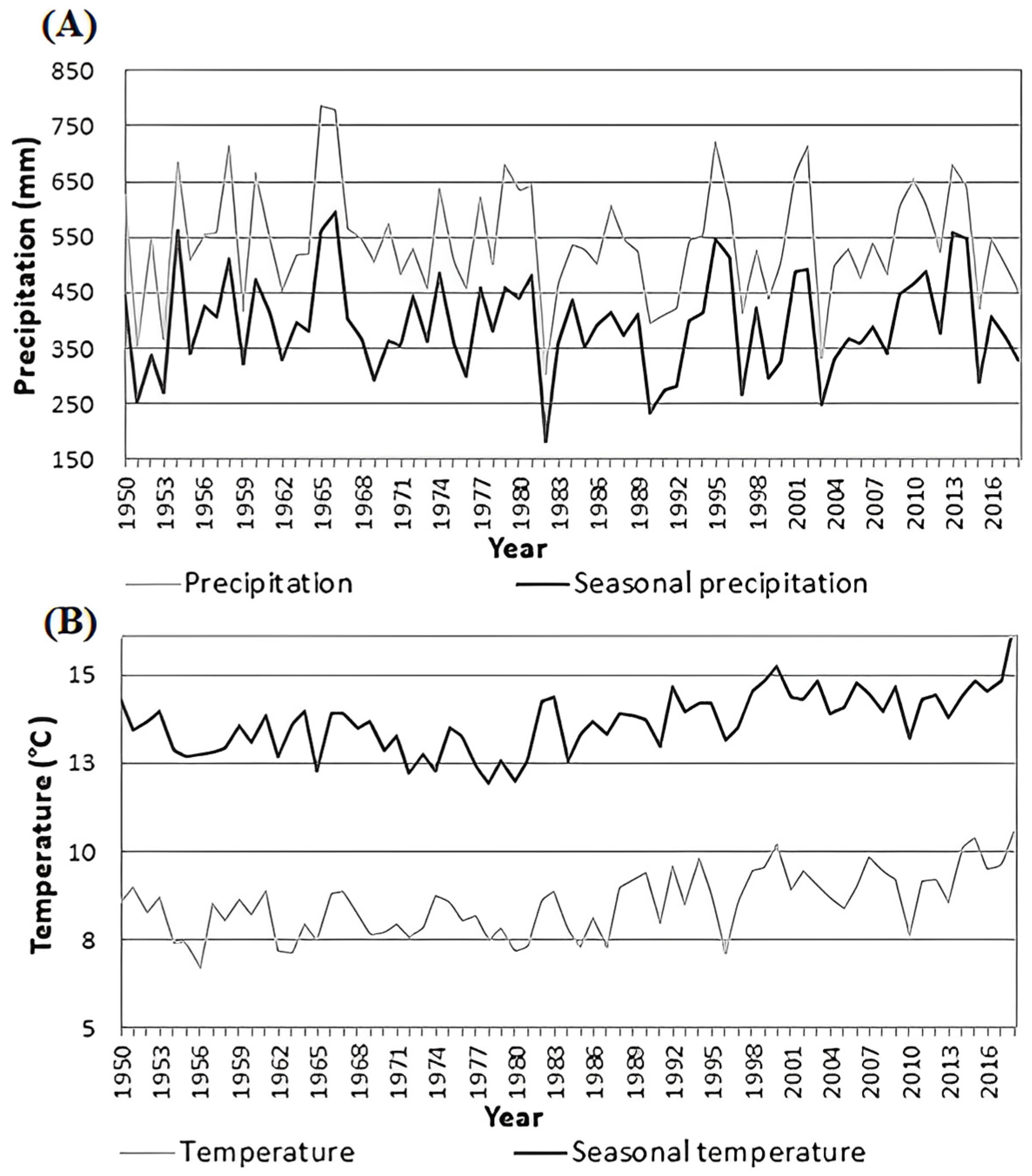
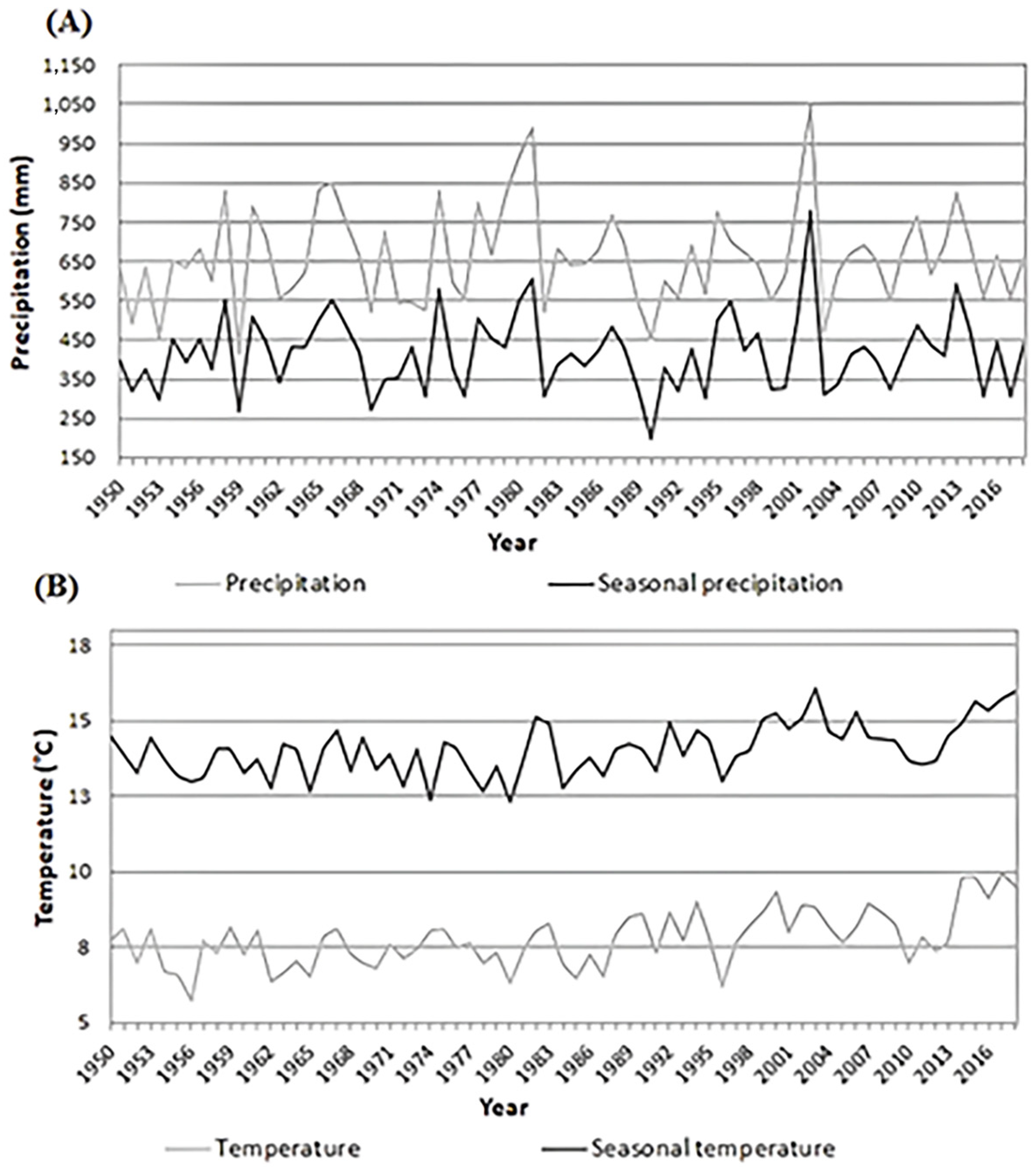
3.1. Dynamics of Radial Growth
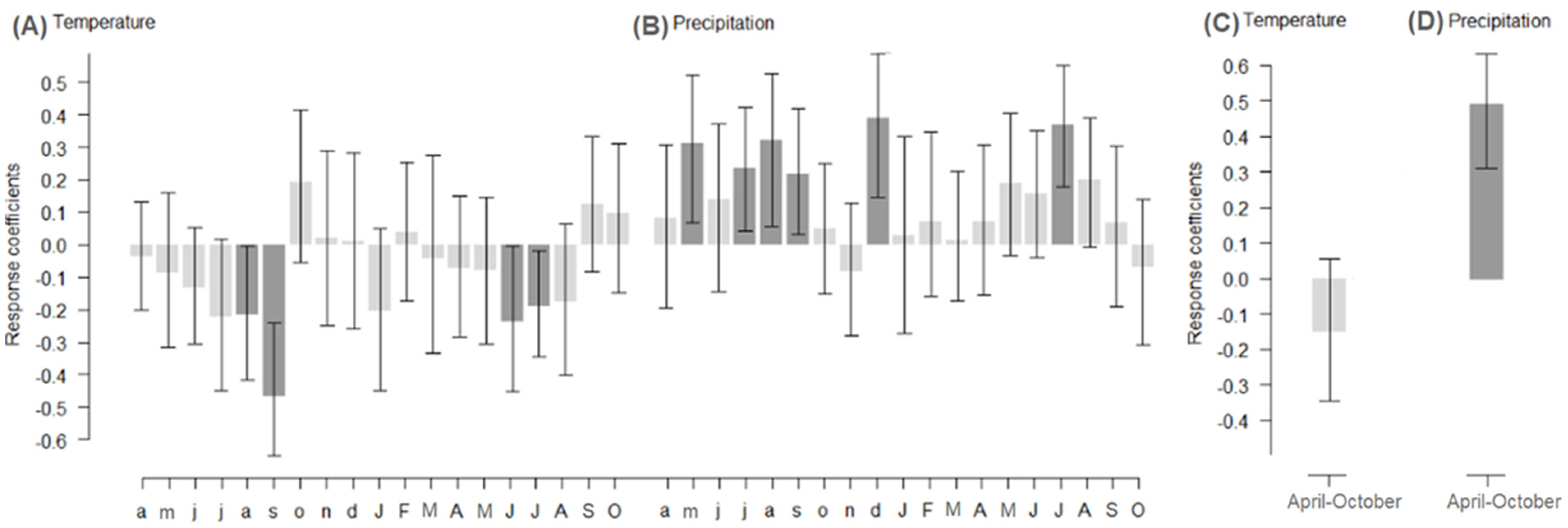
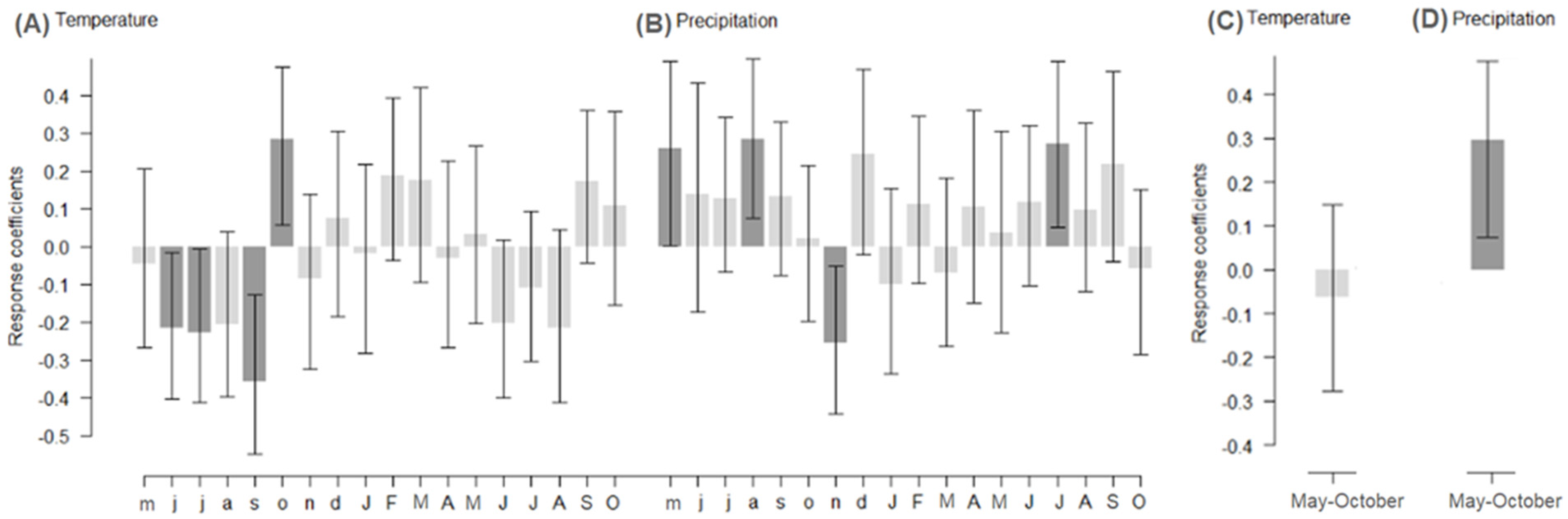
3.2. Effect of Climate on Radial Growth
4. Discussion
4.1. Effect of Climate on Radial Growth
4.2. Dynamics of Radial Growth and Pointer Years
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, L.; Chen, Z.; Olsson, L.; Grahn, T.; Karlsson, B.; Wu, H.X.; Lundqvist, S.O.; García-Gil, M.R. Effect of Number of Annual Rings and Tree Ages on Genomic Predictive Ability for Solid Wood Properties of Norway Spruce. BMC Genom. 2020, 21, 323. [Google Scholar] [CrossRef] [Green Version]
- Rybníček, M.; Čermák, P.; Žid, T.; Kolář, T. Radial Growth and Health Condition of Norway Spruce (Picea abies (L.) Karst.) Stands in Relation to Climate (Silesian Beskids, Czech Republic). Geochronometria 2010, 36, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of Drought Tolerance in Major European Temperate Forest Trees: Climatic Drivers and Levels of Variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- Sidor, C.G.; Popa, I.; Vlad, R.; Cherubini, P. Different Tree-Ring Responses of Norway Spruce to Air Temperature across an Altitudinal Gradient in the Eastern Carpathians (Romania). Trees—Struct. Funct. 2015, 29, 985–997. [Google Scholar] [CrossRef]
- Rybníček, M.; Čermák, P.; Kolář, T.; Žid, T. Growth Responses of Norway Spruce (Picea abies (L.) Karst.) to the Climate in the South-Eastern Part of the Ceskomoravská Upland (Czech Republic). Geochronometria 2012, 39, 149–157. [Google Scholar] [CrossRef]
- Remeš, J.; Pulkrab, K.; Bílek, L.; Podrázský, V. Economic and Production Effiect of Tree Species Change as a Result of Adaptation to Climate Change. Forests 2020, 11, 431. [Google Scholar] [CrossRef] [Green Version]
- Putalová, T.; Vacek, Z.; Vacek, S.; Štefančík, I.; Bulušek, D.; Král, J. Tree-Ring Widths as an Indicator of Air Pollution Stress and Climate Conditions in Different Norway Spruce Forest Stands in the Krkonoše Mts. Cent. Eur. For. J. 2019, 65, 21–33. [Google Scholar] [CrossRef]
- Dudík, R.; Palátová, P.; Jarský, V. Restoration of Declining Spruce Stands in the Czech Republic: A Bioeconomic View on Use of Silver Birch in Case of Small Forest Owners. Austrian J. For. Sci. 2021, 4, 375–394. [Google Scholar]
- Toth, D.; Maitah, M.; Maitah, K.; Jarolínová, V. The Impacts of Calamity Logging on the Development of Spruce Wood Prices in Czech Forestry. Forests 2020, 11, 283. [Google Scholar] [CrossRef] [Green Version]
- Borůvka, L.; Podrázský, V.; Mládková, L.; Kuneš, I.; Drábek, O. Some Approaches to the Research of Forest Soils Affected by Acidification in the Czech Republic. Soil Sci. Plant Nutr. 2005, 51, 745–749. [Google Scholar] [CrossRef]
- Rocha, E.; Gunnarson, B.; Björklund, J.; Zhang, P.; Linderholm, H.W. A Norway Spruce Tree-Ring Width Chronology for the Common Era from the Central Scandinavian Mountains. Dendrochronologia 2021, 70, 125896. [Google Scholar] [CrossRef]
- Koprowski, M.; Zielski, A. Dendrochronology of Norway Spruce (Picea abies (L.) Karst.) from Two Range Centres in Lowland Poland. Trees—Struct. Funct. 2006, 20, 383–390. [Google Scholar] [CrossRef]
- Pichler, P.; Oberhuber, W. Radial Growth Response of Coniferous Forest Trees in an Inner Alpine Environment to Heat-Wave in 2003. For. Ecol. Manag. 2007, 242, 688–699. [Google Scholar] [CrossRef]
- Tumajer, J.; Altman, J.; Štěpánek, P.; Treml, V.; Doležal, J.; Cienciala, E. Increasing Moisture Limitation of Norway Spruce in Central Europe Revealed by Forward Modelling of Tree Growth in Tree-Ring Network. Agric. For. Meteorol. 2017, 247, 56–64. [Google Scholar] [CrossRef]
- D’Andrea, G.; Šimůnek, V.; Castellaneta, M.; Vacek, Z.; Vacek, S.; Pericolo, O.; Zito, R.G.; Ripullone, F. Mismatch between Annual Tree-Ring Width Growth and NDVI Index in Norway Spruce Stands of Central Europe. Forests 2022, 13, 1417. [Google Scholar] [CrossRef]
- Köppen, W. Das Geographische System Der Klimate, Handbuch Der Klimatologie; Gebrüder Borntraeger: Berlin, Germany, 1936. [Google Scholar]
- Pekařová, K. Dynamics of Threatened Species of the Family Rarutnculaceae on the Steppe Localities of Doutnáč Hill in the National Natural Reserve of Karlštejn. Sci. Agric. Bohem. 2007, 3, 24–33. [Google Scholar]
- Vacek, Z.; Vacek, S.; Bílek, L.; Král, J.; Ulbrichová, I.; Simon, J.; Bulušek, D. Impact of Applied Silvicultural Systems on Spatial Pattern of Hornbeam-Oak Forests. Cent. Eur. For. J. 2018, 64, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Rainho, A. Report on the Spot Expert Appraisal of the Karlštejn National Natural Reserve; Council of Europe: Strasbourg, France, 2020; p. 16. [Google Scholar]
- Vacek, S.; Vacek, Z.; Ulbrichová, I.; Bulušek, D.; Prokupková, A.; Král, J.; Vančura, K. Biodiversity Dynamics of Differently Managed Lowland Forests Left to Spontaneous Development in Central Europe. Austrian J. For. Sci. 2019, 136, 249–282. [Google Scholar]
- Tolasz, R.; Míková, T.; Valeriánová, A.; Voženílek, V. Climate Atlas of Czech; Czech Hydrometeorological Institute and Palacký University: Olomouc, Czech Republic, 2007; p. 255. [Google Scholar]
- Šindelář, J.; Frýdl, J.; Novotný, P.; Čáp, J. Silver Fir (Abies alba Mill.) in Limiting Ecological Conditions. In Communicationes Instituti Forestalis Bohemicae; Forestry and Game Management Research Institute: Jíloviště, Czech Republic, 2008; Volume 24, pp. 67–79. ISBN 9788086461922. [Google Scholar]
- Viewegh, J.; Kusbach, A.; Mikeska, M. Czech Forest Ecosystem Classification. J. For. Sci. 2003, 49, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Bílek, L.; Remeš, J.; Zahradník, D. Natural Regeneration of Senescent Even-Aged Beech (Fagus sylvatica L.) Stands under the Conditions of Central Bohemia. J. For. Sci. Sci. 2009, 55, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Podrázský, V.; Remeš, J.; Ulbrichová, I. Vývoj Půdního Chemizmu ve Smrkových Lesních Ekosystémech Na Území ŠLP Kostelec Nad Černými Lesy. Zprávy Lesn. Výzkumu 2005, 50, 198–201. [Google Scholar]
- Remeš, J.; Podrázský, V.V. Fertilization of Spruce Monocultures in the Territory of Training Forest Enterprise in Kostelec Nad Černými Lesy. J. For. Sci. 2006, 52, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Shiyatov, S.G.; Mazepa, V.S.; Ecology, A.; Branch, U. Methods of Dendrochronology Applications; Cook, E.R., Kairiukstis, L.A., Eds.; Tree-Ring Laboratory, Lamont-Ooherty Geological Observatory, Columbia University: New York, NY, USA, 1990; ISBN 9789048140602. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968; p. 110. [Google Scholar]
- Rinntech. TSAP-Win: Time Series Analysis and Presentation for Dendrochronology and Related Applications; Rinntech: Heidelberg, Germany, 2003; Available online: http://www.rimatech.com (accessed on 20 June 2022).
- Larsson, L.A. Cybis Elektronik & Data AB, Saltsjöbaden, Sweden. 2010. Available online: http://www.cybis.se/ (accessed on 20 June 2012).
- Bunn, A.G. A Dendrochronology Program Library in R (DplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. Dendroclimatic Calibration in R: The BootRes Package for Response and Correlation Function Analysis. Dendrochronologia 2013, 31, 68–74. [Google Scholar] [CrossRef]
- Gonzalez, I.G. Weiser: A Computer Program to Identify Event and Pointer Years in Dendrochronological Series. Dendrochronologia 2001, 19, 239–244. [Google Scholar]
- Schweingrub, F.H.; Eckstein, D.; Serre-Bachet, F.; Braker, O.U. Identification, Presentation and Interpretation of Event Years and Pointer Years in Dendrochronology. Dendrochronologia 1990, 8, 9–38. [Google Scholar]
- Rybníček, M.; Čermák, P.; Žid, T.; Kolář, T. Growth Responses of Picea Abies to Climate in the Central Part of the Českomoravská Upland (Czech Republic). Dendrobiology 2012, 68, 21–30. [Google Scholar]
- Kolář, T.; Čermák, P.; Trnka, M.; Koňasová, E.; Sochová, I.; Rybníček, M. Dendroclimatic Study of a Mixed Spruce-Fir-Beech Forest in the Czech Republic. Les/Wood 2020, 69, 21–32. [Google Scholar] [CrossRef]
- Rybníček, M.; Kolář, T.; Čermák, P.; Tomáš, Ž.I.D.; Hadaš, P. Dendrochronological Analysis and Habitual Stress Diagnostic Assessment of Norway Spruce (Picea abies) Stands in the Drahany Highlands. Wood Res. 2012, 57, 189–206. [Google Scholar]
- Affolter, P.; Büntgen, U.; Esper, J.; Rigling, A.; Weber, P.; Luterbacher, J.; Frank, D. Inner Alpine Conifer Response to 20th Century Drought Swings. Eur. J. For. Res. 2010, 129, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Aakala, T.; Kuuluvainen, T. Summer Droughts Depress Radial Growth of Picea Abies in Pristine Taiga of the Arkhangelsk Province, Northwestern Russia. Dendrochronologia 2011, 29, 67–75. [Google Scholar] [CrossRef]
- Bouriaud, O.; Popa, I. Comparative Dendroclimatic Study of Scots Pine, Norway Spruce, and Silver Fir in the Vrancea Range, Eastern Carpathian Mountains. Trees—Struct. Funct. 2009, 23, 95–106. [Google Scholar] [CrossRef]
- Andreassen, K.; Solberg, S.; Tveito, O.E.; Lystad, S.L. Regional Differences in Climatic Responses of Norway Spruce (Picea abies L. Karst) Growth in Norway. For. Ecol. Manag. 2006, 222, 211–221. [Google Scholar] [CrossRef]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced Growth of Alaskan White Spruce in the Twentieth Century from Temperature-Induced Drought Stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- D’Arrigo, R.D.; Kaufmann, R.K.; Davi, N.; Jacoby, G.C.; Laskowski, C.; Myneni, R.B.; Cherubini, P. Thresholds for Warming-Induced Growth Decline at Elevational Tree Line in the Yukon Territory, Canada. Glob. Biogeochem. Cycles 2004, 18, 1–7. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Griesbauer, H.P.; Scott Green, D. Growth Responses of Three Coexisting Conifer Species to Climate across Wide Geographic and Climate Ranges in Yukon and British Columbia. For. Ecol. Manag. 2010, 259, 514–523. [Google Scholar] [CrossRef]
- Grabařová, S.; Martinková, M. Changes of Norway Spruce (Picea abies/L./Karst.) Growth Characteristics under the Impact of Drought. Ekológia 2000, 19 (Suppl. 1), 81–103. [Google Scholar]
- Grabařová, S.; Martinková, M. Changes in Mineral Nutrition of Norway Spruce (Picea abies/L./Karst.) under the Impact of Drought. Ekológia 2001, 20, 46–60. [Google Scholar]
- Kozlowski, A.T.T.; Pallardy, S.G. Acclimation and Adaptive Responses of Woody Plants to Environmental Stresses Published by: Springer on Behalf of New York Botanical Garden Press Acclimation and Adaptive Responses of Woody Plants to Environmental Stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Rita, A.; Camarero, J.J.; Nolè, A.; Borghetti, M.; Brunetti, M.; Pergola, N.; Serio, C.; Vicente-Serrano, S.M.; Tramutoli, V.; Ripullone, F. The Impact of Drought Spells on Forests Depends on Site Conditions: The Case of 2017 Summer Heat Wave in Southern Europe. Glob. Chang. Biol. 2020, 26, 851–863. [Google Scholar] [CrossRef]
- Desplanque, C.; Rolland, C.; Schweingruber, F.H. Influence of Species and Abiotic Factors on Extreme Tree Ring Modulation: Picea Abies and Abies Alba in Tarentaise and Maurienne (French alps). Trees—Struct. Funct. 1999, 13, 218–227. [Google Scholar] [CrossRef]
- Čejková, A.; Čejková, A.; Kolář, T. Extreme Radial Growth Reaction of Norway Spruce Along An Altitudinal Gradient in the Šumava Mountains. Geochronometria 2009, 33, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.J.S.; Hopfmueller, M. Dendrochronological Investigations of Norway Spruce along an Elevational Transect in the Bavarian Forest, Germany. Dendrochronologia 2001, 19, 925–936. [Google Scholar]
- Netherer, S.; Panassiti, B.; Pennerstorfer, J.; Matthews, B. Acute Drought Is an Important Driver of Bark Beetle Infestation in Austrian Norway Spruce Stands. Front. For. Glob. Chang. 2019, 2, 39. [Google Scholar] [CrossRef] [Green Version]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating Outbreak of Bark Beetles in the Czech Republic: Drivers, Impacts, and Management Implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Netherer, S.; Kandasamy, D.; Jirosová, A.; Kalinová, B.; Schebeck, M.; Schlyter, F. Interactions among Norway Spruce, the Bark Beetle Ips Typographus and Its Fungal Symbionts in Times of Drought. J. Pest Sci. 2021, 94, 591–614. [Google Scholar] [CrossRef] [PubMed]
- Bryk, M.; Kołodziej, B.; Pliszka, R. Changes of Norway Spruce Health in the Białowieża Forest (CE Europe) in 2013–2019 during a Bark Beetle Infestation, Studied with Landsat Imagery. Forests 2021, 12, 34. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Boratyński, A.; Bugala, W. Biology and Ecology of Norway Spruce; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea Abies in Europe: Distribution, Habitat, Usage and Threats; European Atlas of Forest Tree Species: Luxembourg, 2016. [Google Scholar]
- Trnka, M.; Hlavinka, P.; Možný, M.; Semerádová, D.; Štěpánek, P.; Balek, J.; Bartošová, L.; Zahradníček, P.; Bláhová, M.; Skalák, P.; et al. Czech Drought Monitor System for Monitoring and Forecasting Agricultural Drought and Drought Impacts. Int. J. Climatol. 2020, 40, 5941–5958. [Google Scholar] [CrossRef]
 ); Climogram for research area of Karlstejn and Cukrak (B); Climogram for study area Kostelec (C)—forested areas are shadowed; the map was created in ArcGIS 10 software version 10.7 (Esri, Redlands, CA, USA); climogram black line describes temperature and grey bars describe precipitation.
); Climogram for research area of Karlstejn and Cukrak (B); Climogram for study area Kostelec (C)—forested areas are shadowed; the map was created in ArcGIS 10 software version 10.7 (Esri, Redlands, CA, USA); climogram black line describes temperature and grey bars describe precipitation.
); Climogram for research area of Karlstejn and Cukrak (B); Climogram for study area Kostelec (C)—forested areas are shadowed; the map was created in ArcGIS 10 software version 10.7 (Esri, Redlands, CA, USA); climogram black line describes temperature and grey bars describe precipitation.
); Climogram for research area of Karlstejn and Cukrak (B); Climogram for study area Kostelec (C)—forested areas are shadowed; the map was created in ArcGIS 10 software version 10.7 (Esri, Redlands, CA, USA); climogram black line describes temperature and grey bars describe precipitation.







| Research Plot | No. Trees | Mean (mm) | Std. | R-Bar | SNR | EPS |
|---|---|---|---|---|---|---|
| Karlstejn | 57 | 1.69 | 0.91 | 0.561 | 71.857 | 0.986 |
| Cukrak | 56 | 1.79 | 0.96 | 0.523 | 61.41 | 0.984 |
| Kostelec | 58 | 1.84 | 0.82 | 0.461 | 49.130 | 0.980 |
| Negative Pointer Year | Abnormal Climatic Characteristics |
|---|---|
| 1964 | Low precipitation in December 1963 and January 1964; subnormal temperature in March. |
| 1976 | Very low precipitation from February to May (especially in February); cold March for Karlstejn and Cukrak. While Kostelec saw very low precipitation in February and April; subnormal temperature in March. |
| 2000 | Very low precipitation in April; supernormal temperature in April and May for Karlstejn and Cukrak. While Kostelec saw very low precipitation in April, subnormal precipitation in June; supernormal temperature from April to June. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Andrea, G.; Šimůnek, V.; Pericolo, O.; Vacek, Z.; Vacek, S.; Corleto, R.; Olejár, L.; Ripullone, F. Growth Response of Norway Spruce (Picea abies [L.] Karst.) in Central Bohemia (Czech Republic) to Climate Change. Forests 2023, 14, 1215. https://doi.org/10.3390/f14061215
D’Andrea G, Šimůnek V, Pericolo O, Vacek Z, Vacek S, Corleto R, Olejár L, Ripullone F. Growth Response of Norway Spruce (Picea abies [L.] Karst.) in Central Bohemia (Czech Republic) to Climate Change. Forests. 2023; 14(6):1215. https://doi.org/10.3390/f14061215
Chicago/Turabian StyleD’Andrea, Giuseppe, Václav Šimůnek, Osvaldo Pericolo, Zdeněk Vacek, Stanislav Vacek, Roberto Corleto, Lukáš Olejár, and Francesco Ripullone. 2023. "Growth Response of Norway Spruce (Picea abies [L.] Karst.) in Central Bohemia (Czech Republic) to Climate Change" Forests 14, no. 6: 1215. https://doi.org/10.3390/f14061215