
Gišogenetic Variation in White-Spruce (Picea glauca (Moench) Voss) Trees of Yukon Beringia, Canada

1
Independent Researcher, Fredericton, NB E3B 4M6, Canada
2
Registered Professional Forester, Victoria, BC V8X 1N6, Canada
3
Independent Researcher, 07973 Greiz, Germany
*
Author to whom correspondence should be addressed.
Forests 2023, 14(4), 787; https://doi.org/10.3390/f14040787
Submission received: 28 February 2023
/
Revised: 30 March 2023
/
Accepted: 31 March 2023
/
Published: 11 April 2023
(This article belongs to the Special Issue Intrinsic Regulation of Diameter Growth in Woody Plants)
Abstract
:Gišogenesis, otherwise known as secondary-xylem development, was investigated in an old-growth upland population of white spruce (Picea glauca (Moench) Voss) trees having morphologically diverse crowns and growing on a south slope north of East Fork Creek bordering never-glaciated Yukon Beringia. After tree felling, trunks were segmented into one-metre lengths. In the laboratory, widths of xylem layers were measured across the four cardinal directions at each height, followed by Pearson’s product momentum correlations to evaluate variation in historical gišogenetic vigour within and between trees. Substantial variation was found, and it cannot readily be explained in terms of differences in extrinsic environment. Physiological differences in intrinsic gišogenetic regulation within a genetically diverse population, comprising both refugia and recent recruits, is proposed as a probable explanation, thus emphasizing the individuality of each tree’s internal control over how it responds to the extrinsic environment. Further investigations within Yukon Beringia may yield insight into evolutionary diversification of gišogenesis.
1. Introduction
In terms of anatomy, chemistry and physics, the properties of wood are variables within individual woody plants, also within and among species [1]. This complexity is magnified by the existence of more than 70,000 species all classified as ‘trees’ [2].
Progress toward understanding intrinsic regulation of meristematic activity has advanced by means of molecular and cell biology research [3], and an ever-increasing number of genetic and hormonal regulators have been characterized (see Discussion). However, the properties of wood change with tree age, and many years are required for the changes to become fully apparent [1]. To investigate this dynamic, one approach is to make direct observations on cambial growth and wood formation in large trees. It is also possible to make indirect deductions, because wood retains a permanent record of changes that occurred as cambial derivatives differentiated into xylem elements [4]. This investigation used both approaches.
Before proceeding, some terminology is clarified. Excepting woody monocots such as bamboos and palms, a circumferential vascular cambium is the progenitor of both phloem and xylem in all tree species. ‘Phloegenesis’ succinctly captures the concept of phloem formation, and a similar term for all aspects of wood formation is needed. ‘Xylogenesis’ has been used but wrongly in this regard because, originally [5] and for decades thereafter, xylogenesis referred specifically to differentiation of cells into tracheary elements. Secondary xylem of no species consists solely of tracheary elements; ray parenchyma are actually the more abundant cell type in most species. Wood was ‘giš’ (rhymes with dish) in the archaic written language of Sumer (the earliest civilization in southern Mesopotamia [6]), so ‘gišogenesis’ is used here to refer to cellular phenomena underlying biogenesis of wood, both during cellular differentiation of radially enlarging and elongating primary-walled cambial derivatives and subsequently during and following completion of their differentiation. ‘Cambial growth’ refers to cell-division activity, the periclinal dividing plane in cambial cells producing derivatives within existing radial files as nascent phloem (‘phloegenesis’) and secondary xylem (‘gišogenesis’) precursors, and the anticlinal plane generating new radial files within the cambial zone [7].
Gišogenesis data are presented here on a white-spruce population north of East Fork Creek bordering the southeastern region of Yukon Beringia (Figure 1). The diversity of crown phenotypes in this population was first noticed in calendar year 1968 [8], followed by investigations to describe them during 2007–2013 [9,10]. In addition, because it was apparent that the plant community had greatly flourished sometime after 1975, the question arose as to whether gišogenesis had been concomitantly altered. In investigating this question, what was unexpectedly observed, as reported here, was within- and between-tree variation in gišogenetic rates during the same calendar-year periods.
The East Fork population was also chosen in anticipation of ongoing research, recognizing that the old growth might hold insight into evolutionary diversification of gišogenesis, an overlooked research realm. After the last glacial maximum (LGM), white spruce re-established in Yukon beginning some 10,000 years ago [11]. However, cryptic refugia probably persisted throughout the LGM and earlier within Eastern Beringia [12], and the diversity at the investigated site is evidence for a refugium. Despite all trees being naturally established, widely spaced with crowns extending from ground level to treetop, a variety of heights, branch-angle and branch-length phenotypes stand together on site [9,10]. Some trees display nearly cylindrical crown silhouettes with flat tops, whereas others appear like bottle brushes, others more like pine trees. A sharply tapered narrow spire silhouette is the usual Yukon crown form characteristic of Picea glauca var. albertiana, and a more spreading but also tapered crown is the usual form of P. glauca var. porsildii, the two commonly recognized Yukon variants of white spruce [10]. The phenotypic heterogeneity so conspicuously present on this site may trace back to ancestors which diversified there as an isolated self-sustaining population [10,13].
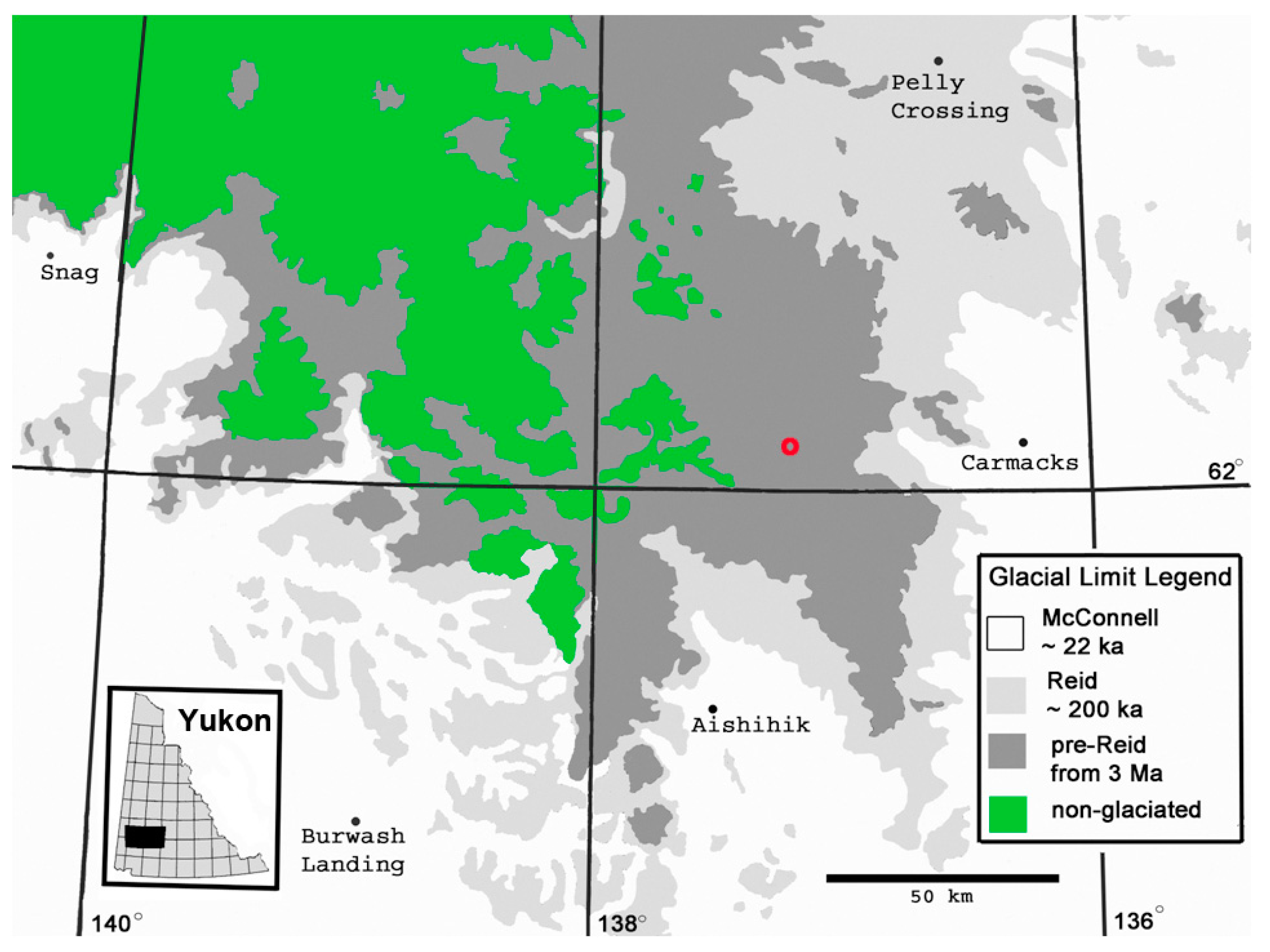
Figure 1.
The red ‘o’ on this map indicates the location of the investigated site in the southeastern Dawson Range, approximately 47 km west of Carmacks, Yukon. This map shows the glacial limits achieved during the Last Glacial Maximum (LGM, or “McConnell”) and those in earlier known ice ages. Some researchers consider any terrain undisturbed by the LGM ice sheet to be within Beringia, whereas others view Beringia as never-glaciated terrain (coloured in green, but see [13]).
Figure 1.
The red ‘o’ on this map indicates the location of the investigated site in the southeastern Dawson Range, approximately 47 km west of Carmacks, Yukon. This map shows the glacial limits achieved during the Last Glacial Maximum (LGM, or “McConnell”) and those in earlier known ice ages. Some researchers consider any terrain undisturbed by the LGM ice sheet to be within Beringia, whereas others view Beringia as never-glaciated terrain (coloured in green, but see [13]).

2. Materials and Methods
2.1. Site Description
The site (Figure 1 and Figure 2), accessible by means of a gravel and mud road (the ‘Mount Nansen road,’ not always traversable) starting from Carmacks, occupies a gentle slope north of East Fork Creek. The geographic centre of its old-growth spruce population is near 62.10556°, −137.1878°.
The site is in the mountains of the southern Dawson Range within the coldest region of North America. By straight line measure, it is 47 km west of Carmacks at 543 m above sea level (asl), 58 km north northeast of Aishihik (914 m asl), 70 km south of Fort Selkirk (458 m asl, record holder of the coldest unofficial North American temperature of −65 °C) and 170 km east of Snag (550 m asl, record holder of the lowest official North American temperature of −62.8 °C). Before clouds arrive overhead, their moisture is mostly removed over distant high mountain regions; thus a semi-arid climate prevails [14].
East Fork Creek flows into Nansen Creek at an elevation of 1180 m asl. Two km upstream, the East Fork valley bottom rises to 1250 m asl. The trees selected for investigation were all above 1300 m asl on a south aspect north of East Fork Creek. In Figure 2A, the visible lower slope trees are near 1240 m asl; those at timberline are near 1400 m asl.
Figure 2.
The investigated site in early September (A–C) and early October (D) 2007. (A) Looking north, the foreground trees are at 1240 m asl and those at highest elevation are near 1400 m asl. The vehicle is 6 m in length. Horizontal distance between the left summit and the highest summit on the right is 1.25 km. (B) The same site looking west with Mount Nansen in the distance and the confluence of East Fork Creek with Nansen Creek the lowest point. (C,D) Closer views of the white spruce trees on site, all widely spaced and full crowned; some lean due to downslope slippage over melting permafrost.
Figure 2.
The investigated site in early September (A–C) and early October (D) 2007. (A) Looking north, the foreground trees are at 1240 m asl and those at highest elevation are near 1400 m asl. The vehicle is 6 m in length. Horizontal distance between the left summit and the highest summit on the right is 1.25 km. (B) The same site looking west with Mount Nansen in the distance and the confluence of East Fork Creek with Nansen Creek the lowest point. (C,D) Closer views of the white spruce trees on site, all widely spaced and full crowned; some lean due to downslope slippage over melting permafrost.

The site’s gentle south aspect formed more than a million years ago (Ma). Rocky elevations above (Figure 2A) are possibly remnants of a deglaciation that occurred 1.6 Ma [15]. The East Fork valley bottom holds glacial drift [16], but none was found at 1300 m asl. Depth to bedrock was not established, due to permafrost.
The on-site community (Figure 2) comprises widely spaced mature white-spruce trees, almost all having a full live crown. Scattered small black-spruce (Picea mariana (Mill.) B.S.P.) trees, many evidently clonal through layering, grow along the East Fork valley bottom, probably derived from a large population on the west side of Nansen Creek. Single individuals of black spruce were encountered but rarely within the East Fork population. Shrubbery is dominantly Betula glandulosa Michx. interspersed with several Salix species. Ground beneath white-spruce crowns has little vegetation; it is probable that some form of allelopathy is operative [17]. Between trees, perennial vascular plants, bryophytes and lichens grow in tight association as a spongy tundra carpet. Inexplicably, no Alnus or tree-sized Betula or Populus species were encountered on site, although they are common in the surrounding region at elevations up to 1100 m asl and exist in other Yukon regions at elevations exceeding 1300 m asl. Some young Populus tremuloides Michx. and P. balsamifera L. trees established near road edges and on placer-mining spoils over the last two decades, but mature specimens were not seen on site. A list of ground flora encountered in the locale is provided in Table S1 from Supplementary Materials. Specimens CANL 124,983–125,017 and CANL 331,528–331,579 were deposited with the National Herbarium of Canada.
In response to gelifraction, cryoplanation and the occasional torrential rain storm, eroded materials have accumulated as slope colluvium [13]. Leaning trunks (Figure 2B,C) indicate that the rooting zone in localized patches of tundra has experienced downward slippage over the permafrost table, but this has occurred only in rare locations.
On-site temperature data were gathered using MadgeTech cryo-temp low temperature units (MadgeTech Inc., Warner, NH, USA) programmed to record temperature hourly. All loggers were pre-calibrated and confirmed accurate within ±1 °C. The collected data revealed that air temperatures paralleled those at the government weather station in Carmacks, similar fluctuations affecting this region’s subdued mountain landscape over an east-west distance of more than 100 km (Figure 3 and Figure 4).
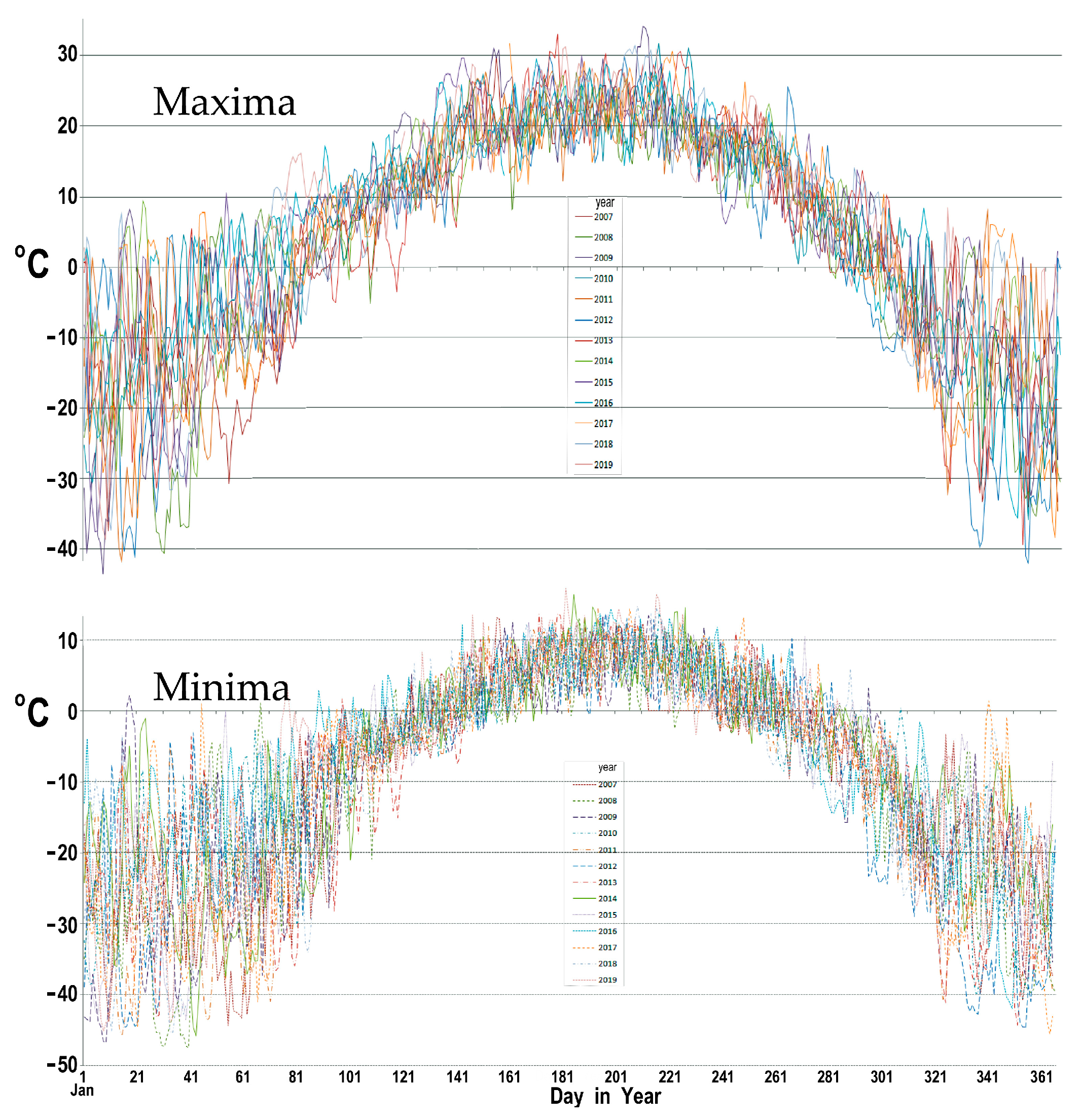
Figure 3.
Mean maxima and minima daily temperatures at Carmacks, Yukon based on 2007 to 2019 data gathered by Environment Canada.
Figure 3.
Mean maxima and minima daily temperatures at Carmacks, Yukon based on 2007 to 2019 data gathered by Environment Canada.

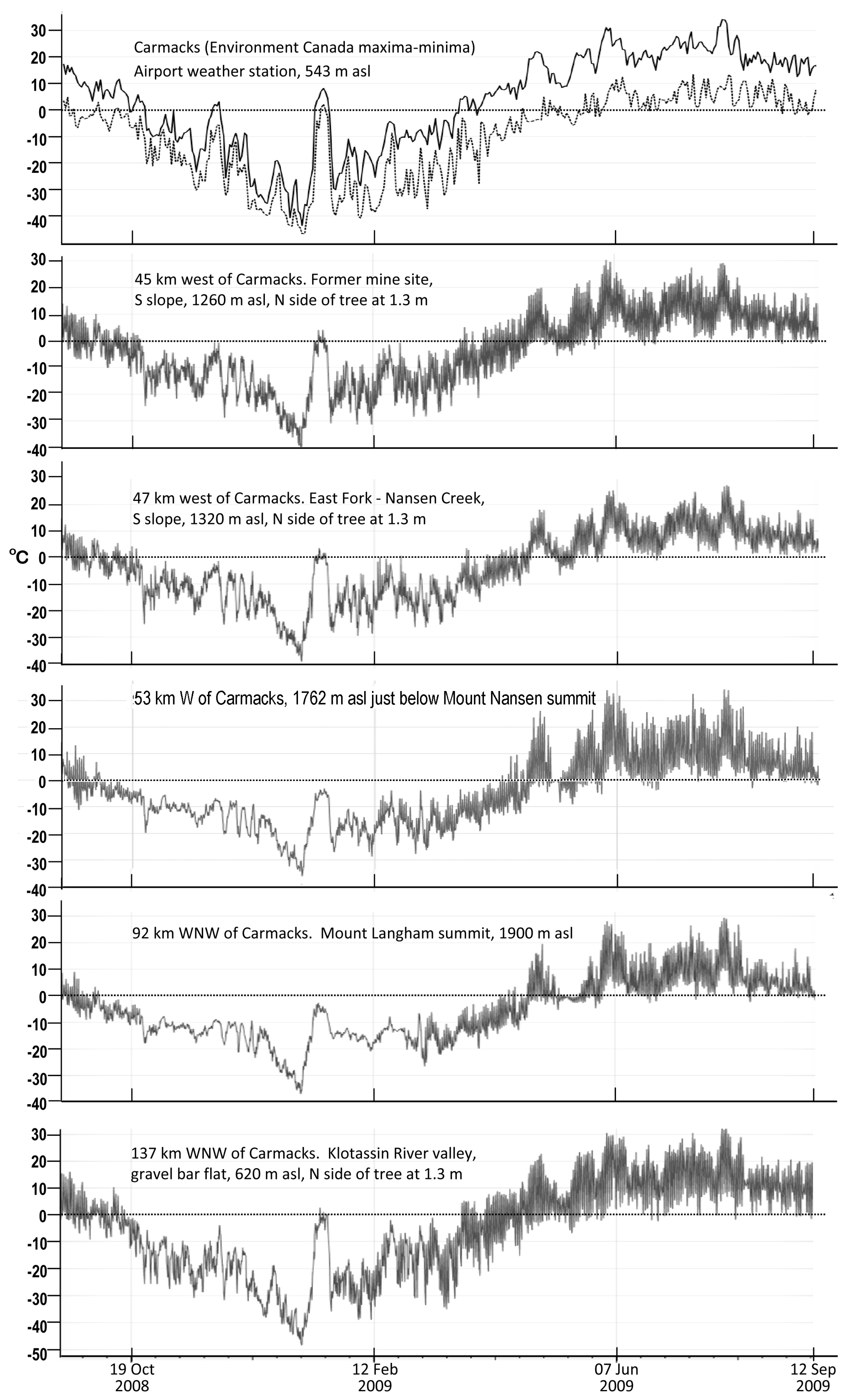
Figure 4.
Temperature data for the East Fork site and locations to east and west for the 2008–2009 calendar years. Field data were logged with MadgeTech cryo-temp low temperature units.
Figure 4.
Temperature data for the East Fork site and locations to east and west for the 2008–2009 calendar years. Field data were logged with MadgeTech cryo-temp low temperature units.

Official maximum and minimum temperatures as recorded daily at Carmacks airport (543 m asl) over a 13-year period indicate that greatest year-to-year variation occurs from October through March (Figure 3 and Figure 4). On the basis of the adiabatic lapse rate for dry air, the East Fork site at 1300 m asl should have air temperatures nearly 7 °C below those recorded at Carmacks. That appeared to be the case from June through September, the months when snow was not present on site during the 2007–2013 calendar years (Figure 5). However, our data from other sites indicated that additional factors influence air temperatures in the region. For example, summertime temperatures logged on the East Fork site and in the Klotassin River valley bottom were very similar in magnitude to those logged on a plateau less than 100 m below the 1827 m asl summit of Mount Nansen and upon the Mount Langham summit (Figure 4). Temperatures on the two mountain tops were slightly above those on the East Fork site during the winter months, probably explainable in terms of greater snow accumulation on the summits and night-time down- slope air flow and valley pooling of cold air.
To gain insight into the thermal environment surrounding the trees on the East Fork site during a growing season, high-resolution MadgeTech cryo-temp data loggers were placed on the ground and at one and two metres above, suspended beneath tree branches (Figure 5A). Additional Hobo UA-002-64 (ver.1.0.8) pendant temperature/light data loggers (Onset Computer Corp., 470 MacArthur Blvd., Bourne, MA, USA) were suspended on the trunk surface at higher heights (Figure 5B).
On-site and Carmacks data mostly varied in parallel (Figure 5A), but they also revealed that Carmacks temperatures do not always provide an accurate impression of extremes experienced on site even after adiabatic lapse rate adjustment.
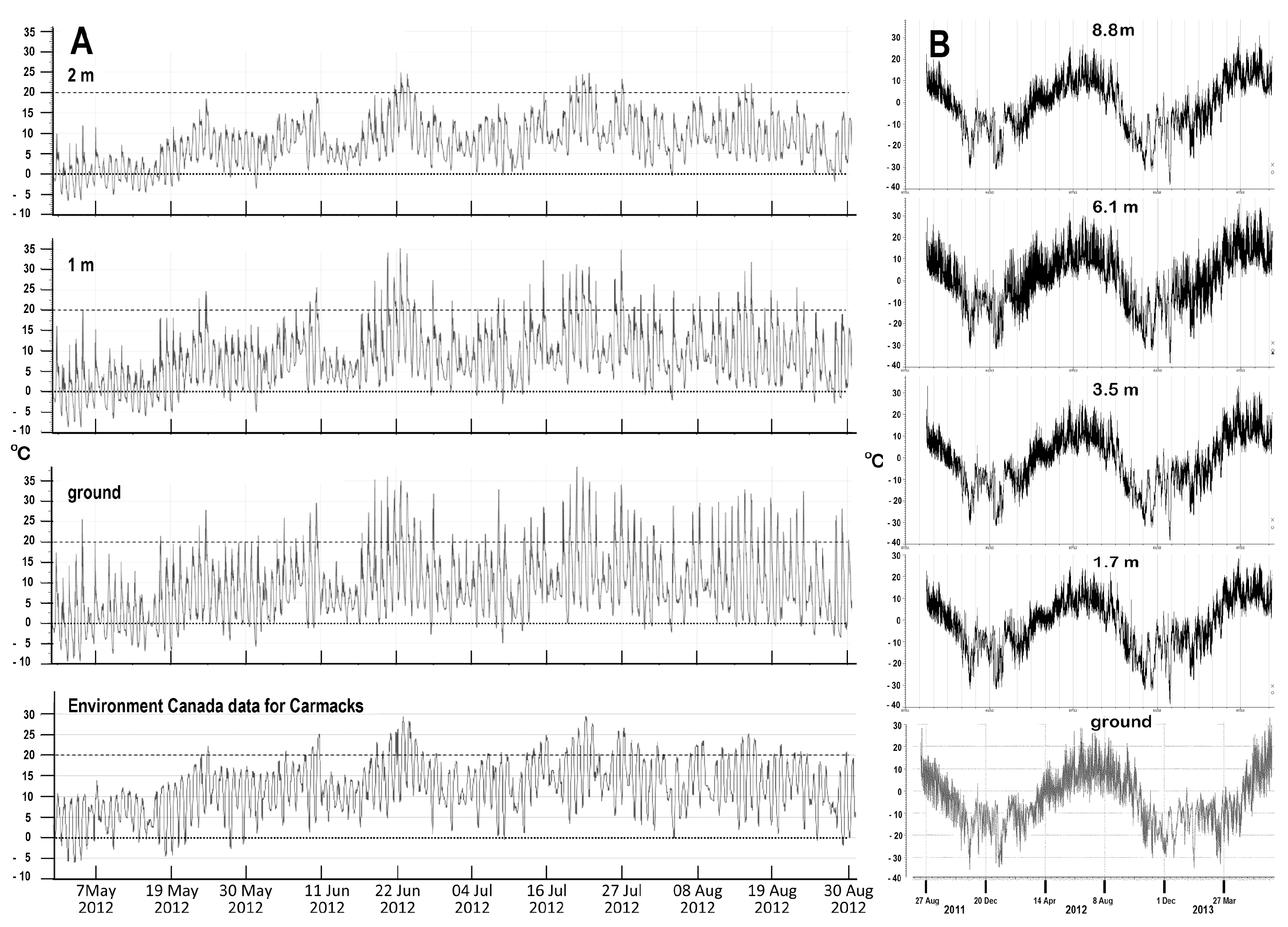
Figure 5.
Temperature data for several trunk heights at one location on the East Fork site. (A) Growing season temperatures at ground level and one and two metres above, compared with official data recorded at Carmacks, during the summer of 2012. (B) Air temperature fluctuations during calendar years 2011–2013 at ground level and four heights above. The slightly warmer ground temperatures during winter are presumably due to snow cover.
Figure 5.
Temperature data for several trunk heights at one location on the East Fork site. (A) Growing season temperatures at ground level and one and two metres above, compared with official data recorded at Carmacks, during the summer of 2012. (B) Air temperature fluctuations during calendar years 2011–2013 at ground level and four heights above. The slightly warmer ground temperatures during winter are presumably due to snow cover.

On-site daytime temperatures hovered around 20 °C from late June to early August, rarely approaching 30 °C. Buds were fully dormant and microscopy confirmed that cambium was entering dormancy in early September. During that summer period, night-time temperatures were commonly below 10 °C (Figure 5A) and often dropped below freezing at and just above ground level (Figure 5A,B). No evidence was seen for freezing damage; growth, flowering and general health all appeared resilient. Plant hardiness on site is clearly adapted such that even tender new growth can survive under daily temperature fluctuations ranging from <0 °C to 30 °C.
Large diameter horizontal roots near the trunk bases of old-growth spruce trees are common above the tundra surface on site. Frost heaving may be part of the explanation, but cross sections of a young and an older root revealed that gišogenesis is strongly in favor of zenith increments (Figure 6A,B). Such eccentricity in root secondary growth is well known, but on this site the different growth rates on opposing sides evidently are emphasized by differences in temperatures. The cold ground presumably impedes cambial growth on the nadir side. Temperatures in the soil remain annually <10 °C, while on and above the tundra surface temperatures can exceed 30 °C in summer (Figure 7). Small- diameter roots are present in the dark brown organic horizon to a depth of 60 cm; fine roots are also present but rarely encountered in the lighter brown volcanic felsic mineral soil below (Figure 6C). In one excavation, a 6-cm long earthworm, possibly Aporrectodea caliginosa, was unexpectedly encountered in the organic horizon [18]. White-spruce roots form associations with both ecto- and endo-mycorrhizae and, although roots were not investigated in that respect, numerous species of fungi were present on site (Table S2).
Figure 6.
(A) A cross-section at one metre from the root collar through two horizontal roots; the smaller root at lower right egresses from the larger and was growing buried within tundra, whereas the larger root was above the tundra surface. The arrows indicate positions of the root centres in their first years. (B) Cross-section of an older horizontal root. The arrow at bottom indicates the centre of the root when it was in its first year. (C) Organic soil overlaying solidly frozen colluvium at 1300 m asl on site. The long white bar at right indicates a residual grey tephra layer deposited more than 1200 years ago [19]. The mattock handle is 90 cm in length.
Figure 6.
(A) A cross-section at one metre from the root collar through two horizontal roots; the smaller root at lower right egresses from the larger and was growing buried within tundra, whereas the larger root was above the tundra surface. The arrows indicate positions of the root centres in their first years. (B) Cross-section of an older horizontal root. The arrow at bottom indicates the centre of the root when it was in its first year. (C) Organic soil overlaying solidly frozen colluvium at 1300 m asl on site. The long white bar at right indicates a residual grey tephra layer deposited more than 1200 years ago [19]. The mattock handle is 90 cm in length.

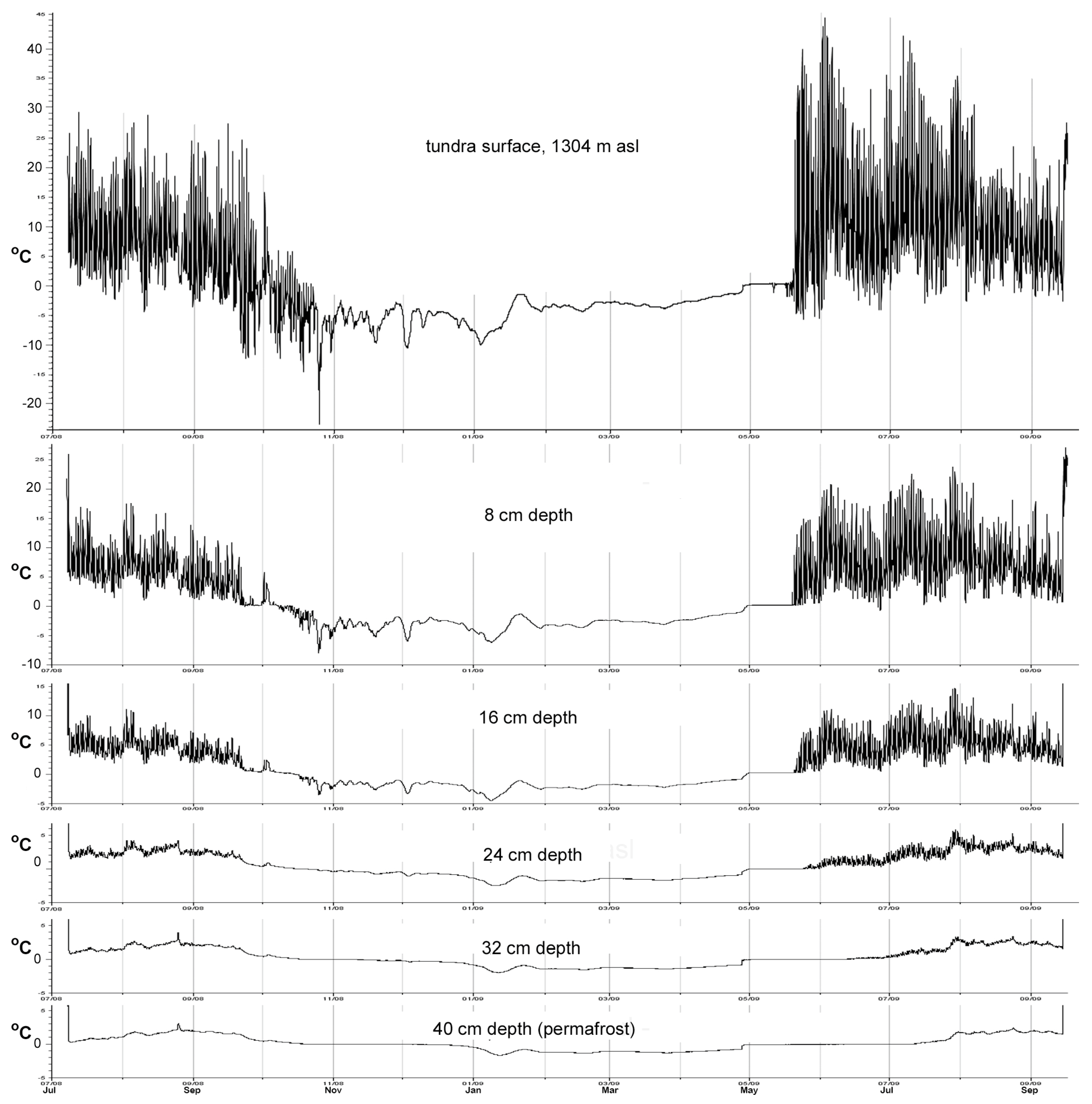
Figure 7 shows how soil temperatures on site varied at several depths, based on July 2008 to September 2009 data. The ground surface remained beneath snow cover from mid-October to mid-May and thereafter warmed up during the June–September months. At depths of 24 cm and deeper beneath the surface, temperatures were above freezing during summer months and very slightly below 0 °C during winter months. Unfrozen soil, whether saturated or not, usually freezes at a temperature several degrees below 0 °C [20]. Thus, it seems probable that cold meltwater at the permafrost–active layer interface is available for upward capillarity and root absorption when air temperatures become conducive to growth.
Soil moisture content below the surface was investigated by collecting random unfrozen soil samples at 10 cm depth on a dry day in early July (Table S2). Moisture content, determined by subtraction of oven-dried from fresh weight, was 81% (std. dev. 11%). The pH of those soils, measured by calibrated pH electrode, ranged from pH 3.26 to pH 5.08.
Figure 7.
July 2008 to September 2009 variation in soil temperature at the surface and five successive depths spaced 8 cm apart, the 40 cm depth being just above the permafrost table. Data were gathered using Hobo UA-002-64 light/temperature data loggers. Light data were used to confirm that the loggers remained buried.
Figure 7.
July 2008 to September 2009 variation in soil temperature at the surface and five successive depths spaced 8 cm apart, the 40 cm depth being just above the permafrost table. Data were gathered using Hobo UA-002-64 light/temperature data loggers. Light data were used to confirm that the loggers remained buried.

The on-site surface contains dry fallen snags (Figure 8A), and some give the impression of having lain on the ground for many years. To investigate whether biotic turnover occurs in the soil, poplar sticks with half lengths painted fluorescent red (for identification), oven dried, and pre-weighed were inserted to half depth into soil in triplicate at random sites in early July 2008 (Figure 8B).
Figure 8.
(A) A fallen snag that has lain on the tundra surface for >30 years. Severely spiral-grained specimens are common on site, but so too are perfectly straight-grained trees. (B) An example of poplar sticks used to investigate biomass turnover from July 2008 to September 2009.
Figure 8.
(A) A fallen snag that has lain on the tundra surface for >30 years. Severely spiral-grained specimens are common on site, but so too are perfectly straight-grained trees. (B) An example of poplar sticks used to investigate biomass turnover from July 2008 to September 2009.

In early September 2009, 274 of those sticks were retrieved, then cleansed of adhering particles, washed, oven dried and reweighed in the laboratory. The upper painted portions remained visibly unaltered over 14-month field exposure. Making the assumption that the entire stick if unpainted and fully inserted would have deteriorated to the same extent as the insert half, the data yielded a mean biomass decomposition of 23.5% (w/w; range 2.6%–75.6%, std. dev. 11.0%). Microbial decomposition is supported by observations on site of mushroom diversity (Figure S1).
Forest fires are very common at lower elevations in this region of Eastern Beringia [21]. However, despite searching for charcoal in more than 40 soil excavations on the East Fork site, none was encountered. This higher elevation taiga site evidently has never experienced fire.
Climate change in recent decades evidently enhanced plant establishment and growth on site. During 1968–1975 investigations, a valley glacier having a surface height of several metres lay over the full length of East Fork Creek throughout summer months. In August, at the height of summer warming, snow and ice remained on northern aspects, and the vertical depth to solid permafrost was <10 cm everywhere. There were no small trees found on site, and dwarf birch and willow shrubs were only knee high and displayed few, if any, green leaves (Figure 9A–C). Excepting one brief examination of a spiral-grained snag (Figure 9A) [4], gišogenesis investigations were not conducted during the 1968–1975 period.
Figure 9.
Early August 1974. Three on-site photographs taken at 1250 m ± 20 m asl; (A) looking west with East Fork Creek in the valley bottom at left; inset: xylem layers in a cross section of a branch from the foreground tree (see [6]). (B) Looking west; (C) looking east; the axe handle = 60 cm.
Figure 9.
Early August 1974. Three on-site photographs taken at 1250 m ± 20 m asl; (A) looking west with East Fork Creek in the valley bottom at left; inset: xylem layers in a cross section of a branch from the foreground tree (see [6]). (B) Looking west; (C) looking east; the axe handle = 60 cm.

Upon returning in early June 2007, the birch-willow shrubbery was densely foliated and had heights ~2 m. The East Fork valley bottom was entirely free of ice. By the end of July, north slopes and mountaintops were also bare of snow, and depth to impenetrable permafrost was ~40 cm. Spruce saplings were present among the old-growth population (Figure 10A) and, based on an approximately 30-year age of the tallest found (Figure 10A,B), there was capability for seedling establishment by the early 1980s. Seedlings at altitudes >1400 m asl were found in calendar year 2011, on sites where trees had not previously existed (Figure 10C,D).
Figure 10.
(A) The tallest (ca. 6 m) sapling found on site in June 2007 is beside the man and was growing at 1254 m asl, 250 m N of East Fork Creek. (B) A view 10 cm above the root collar of the tree shown in (A); its age is estimated at ~30 years. (C) A 23-cm-tall seedling found at 1470 m asl, and (D) a section near the seedling’s root collar showing 9 xylem layers (no chemical fixative; the orange zone comprises dead cambial derivatives ). Stained with 0.05% (w/v) toluidine blue O (see [22]).
Figure 10.
(A) The tallest (ca. 6 m) sapling found on site in June 2007 is beside the man and was growing at 1254 m asl, 250 m N of East Fork Creek. (B) A view 10 cm above the root collar of the tree shown in (A); its age is estimated at ~30 years. (C) A 23-cm-tall seedling found at 1470 m asl, and (D) a section near the seedling’s root collar showing 9 xylem layers (no chemical fixative; the orange zone comprises dead cambial derivatives ). Stained with 0.05% (w/v) toluidine blue O (see [22]).

2.2. Tree Investigations
Six mature white-spruce trees within the East Fork population were selected for felling (Figure 11). The tallest (16.2 m) was felled 12 June 2007, the other five in late June and early July 2008. Hereafter, these individuals are referred to by their elevations, e.g., the one at 1311 m asl is ‘tree 1311’.
Figure 11.
The six investigated trees photographed just before being felled in 2007 (1316 m) and 2008 (the other five trees). The 8 m South radius of tree 1311 in cross section, and the tree top and stage of budbreak of tree 1316 are shown at left and right, respectively, of the top of tree 1316.
Figure 11.
The six investigated trees photographed just before being felled in 2007 (1316 m) and 2008 (the other five trees). The 8 m South radius of tree 1311 in cross section, and the tree top and stage of budbreak of tree 1316 are shown at left and right, respectively, of the top of tree 1316.

Altitudes and geographic coordinates of the six (Table 1) were obtained by hand-held gps, using World Geodetic System (WGS-84) coordinates with the Wide Area Augmentation System (WAAS). Positional errors displayed by the gps unit were from 10 to 20 m; however, by revisiting sites over three successive years using the identical system, accurate return ±3 m was possible.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Altitudes, trunk lengths and locations of the six investigated trees.
| Altitude | Height | Coordinates | ||
|---|---|---|---|---|
| (m asl) | (m) | North | West | |
| 1311 | 8.4 | 62.1089 | −137.19905 | |
| 1316 | 16.2 | 62.1021 | −137.18319 | |
| 1320 | 10.2 | 62.1026 | −137.18467 | |
| 1326 | 9.4 | 62.1033 | −137.18650 | |
| 1328 | 9.5 | 62.1032 | −137.18633 | |
| 1329 | 6.8 | 62.1036 | −137.18764 |
Before a tree was felled, true North (magnetic declination 18.5° E) was determined by compass and a mark made on the north side of its trunk. The felled trunk was de-branched then cut transversely into segments 1.00 m ± 0.05 m in length. The North direction was marked on each segment’s lower end, after which a disk approximately 5 cm in trunk axial length was cut from that end for transfer to the laboratory.
The six trees were felled at heights approximately 30 cm above ground level; thus 30 cm should be added to all stated heights (Table 1), and the “1 m” trunk-height position is actually ~1.3 m, or breast height. The full ages of the felled trees could not be estimated. Yukon government classifies this upland as non-forested, and no estimates could be found on the number of years it may take, following germination, for a seedling to reach a height of 30 cm. During calendar years 2007/2008, some on-site “seedlings” were determined to be ~10 years old (Figure 10C,D). However, it would be assumptive to think a similar rate of height growth occurred when the six investigated trees were seedlings. In the laboratory, the upper, unmarked surface of each trunk disk was power planed, sanded using successively finer grades of Siafast (60, 80, 120, 220 grit) abrasive discs, followed by Norton SandWet (Norton | Saint-Gobain Abrasives, Worcester, MA, USA) (400, 600 grit) sandpaper, polishing with 1500 and 2000 grit waterproof automotive sandpaper and wet finishing with 7940 Siaair 2000 and 4000 grit pads (SMS Distributors, East Falmouth, MA, USA). Straight pencil lines were drawn at right angles across the wood surface from the pith to cambium, to indicate the four cardinal-direction radii.
Immediately before scanning a radius across a disk surface using WinDendro v. 2001b (Regent Instruments Inc., Quebec, Canada) in conjunction with a 1680 Epson Expression scanner (800 dpi) (Seiko Epson Co., Suwa, Nagano, Japan) the disk was sprayed with water to enhance resolution. Distances were measured from one latewood - earlywood boundary to the next (hereafter referred to as ‘ring width’) from vascular cambium to pith. The incompletely formed annual ring in process of development at time of tree felling was not measured.
A precision stage micrometer (Wild, Heerbrugg, Switzerland): 100 divisions of 0.10 ± 0.02 mm per cm) was used for calibration, and WinDendro precision was determined to be 25 ± 10 µm. When compared with higher resolution microscopy measurements, WinDendro had a consistent bias toward underestimation, but the difference was so slight that no correction was made. In view of the precision of both methods, replicate estimates of ring widths were not made.
Three of the six trees had decay with ant galleries in the lowermost (0 m height) disk, making data over all four cardinal directions at that 30-cm height unobtainable. However, the wood in all six trees was sufficiently sound at 1-metre and higher trunk heights for complete data sets to be obtained across all four cardinal directions. WinDendro was unable to resolve all ring widths at 7 m, 8 m and treetop of tree 1311 (see Figure 11), also at 6 m of tree 1316. Unresolvable sectors were sawn to a size suited for the inverted microscope stage, and higher precision (10 µm ± 1 µm) light-microscope measurements on those radii were made using a calibrated eyepiece micrometer.
On the assumption that each layer of xylem corresponded to one calendar year of gišogenesis, each ring-width measurement in a radial series was assigned a calendar year, proceeding inward from cambium to pith. Agreement in numbers of layers over the four radii verified full circumferential rings. No transformations were done before comparing the same calendar-year periods over a radial series for similarity using the Pearson product–moment–correlation coefficient (−1 ≤ r ≤ +1). The four cardinal directions were compared against each other at each trunk height. Within-tree trunk heights were investigated by comparing the same cardinal direction. Between-tree comparisons were based on the mean ring widths of the four cardinal directions for each year of secondary xylem. In Pearson’s correlations, an r value approaching +1 over the same period indicates that ring widths (or mean ring widths in the case of between-tree comparisons) in two chronological series were similarly in the process of increasing or decreasing or remaining constant, whereas an r value approaching −1 indicates that radial width increments were trending (or sloping) with the passage of years in opposite directions. In other words, an r value of +1 indicates a perfect direct correlation, an r value of −1 a perfect inverse correlation and an r value of 0 no correlation. An r value of +1 does not require that for every unit increase (or decrease) in one chronological series there is exactly a unit increase (or decrease) in the other’s corresponding series, rather, it merely requires that the two have similar slopes relative to earlier and later years.
On 12 June 2007, tree 1316 displayed early bud swelling (Figure 10), enabling investigation of cambial reactivation in its trunk. The midpoint of the south-facing side of each 1-metre segment was sampled (Figure 11). Lacking fixation chemicals, specimens were preserved in 1:1 (v/v) 70% isopropanol: white vinegar until transported to the laboratory for light and scanning electron microscopy (SEM). Sections were hand cut and stained using 0.05% toluidine blue O for 60 s, then rinsed and examined under brightfield illumination as described previously [22]. Radial sections were viewed by differential interference contrast microscopy and, if bordered pits were in the process of formation in primary- walled ‘RE’ cambial derivatives, those enlarged cells were considered in the process of secondary-wall formation (‘SL’ zone, see [23]). SEM was performed as earlier described [23].
Using a manual increment borer, cores from trunks of a number of trees on site were extracted at 1.3 m height. In the laboratory, the cores were razor shaved, stained 30 s with pyronin Y (Sigma-Aldrich Inc., St. Louis, MO, USA) in water (10 mg/mL) to enhance latewood–earlywood contrast, and imaged using an inverted light microscope (Figure 12). Ten-year measurements to the nearest tenth millimetre were made from 1967 to 2007 to determine rates of gišogenesis.
Figure 12.
(A) Part of a white-spruce core before and after staining with pyronin Y. Examples of variation in cores obtained at 1.3 m trunk height from randomly sampled white-spruce trees. (B) Breast-height cores extracted in 2008 from trunks of 8 trees, showing the latewood–earlywood boundary as a black horizontal dash between the 1980 and 1981 growth layers.
Figure 12.
(A) Part of a white-spruce core before and after staining with pyronin Y. Examples of variation in cores obtained at 1.3 m trunk height from randomly sampled white-spruce trees. (B) Breast-height cores extracted in 2008 from trunks of 8 trees, showing the latewood–earlywood boundary as a black horizontal dash between the 1980 and 1981 growth layers.

3. Results
3.1. Gišogenesis Based on Raw Ring-Width Measurements
Within-trunk variation between cardinal directions displaying maximum overall gišogenesis varied with trunk height (Table 2). The north side was very slightly more favored over the south side, but east and west sides were equivalently favored and probably not significantly different from north and south.
Table 2.
The cardinal direction having greatest radial width from pith to cambium at each trunk height of each of the six trees.
Table 2.
The cardinal direction having greatest radial width from pith to cambium at each trunk height of each of the six trees.
| Height | Tree | |||||
|---|---|---|---|---|---|---|
| (m) | 1311 | 1316 | 1320 | 1326 | 1328 | 1329 |
| 16 | E | |||||
| 15 | N | |||||
| 14 | N | |||||
| 13 | N | |||||
| 12 | S | |||||
| 11 | S | |||||
| 10 | W | E | ||||
| 9 | N | E | * | N | ||
| 8 | W | N | S | E | N | |
| 7 | S | N | W | N | N | |
| 6 | S | N | N | W | S | S |
| 5 | E | E | N | S | S | * |
| 4 | S | E | N | E | W | S |
| 3 | W | E | E | N | W | S |
| 2 | W | N | W | W | N | S |
| 1 | E | E | S | E | N | N |
| 0 | S | W | S | W | N | N |
An asterisk (*) indicates that the direction could not be resolved at that position.
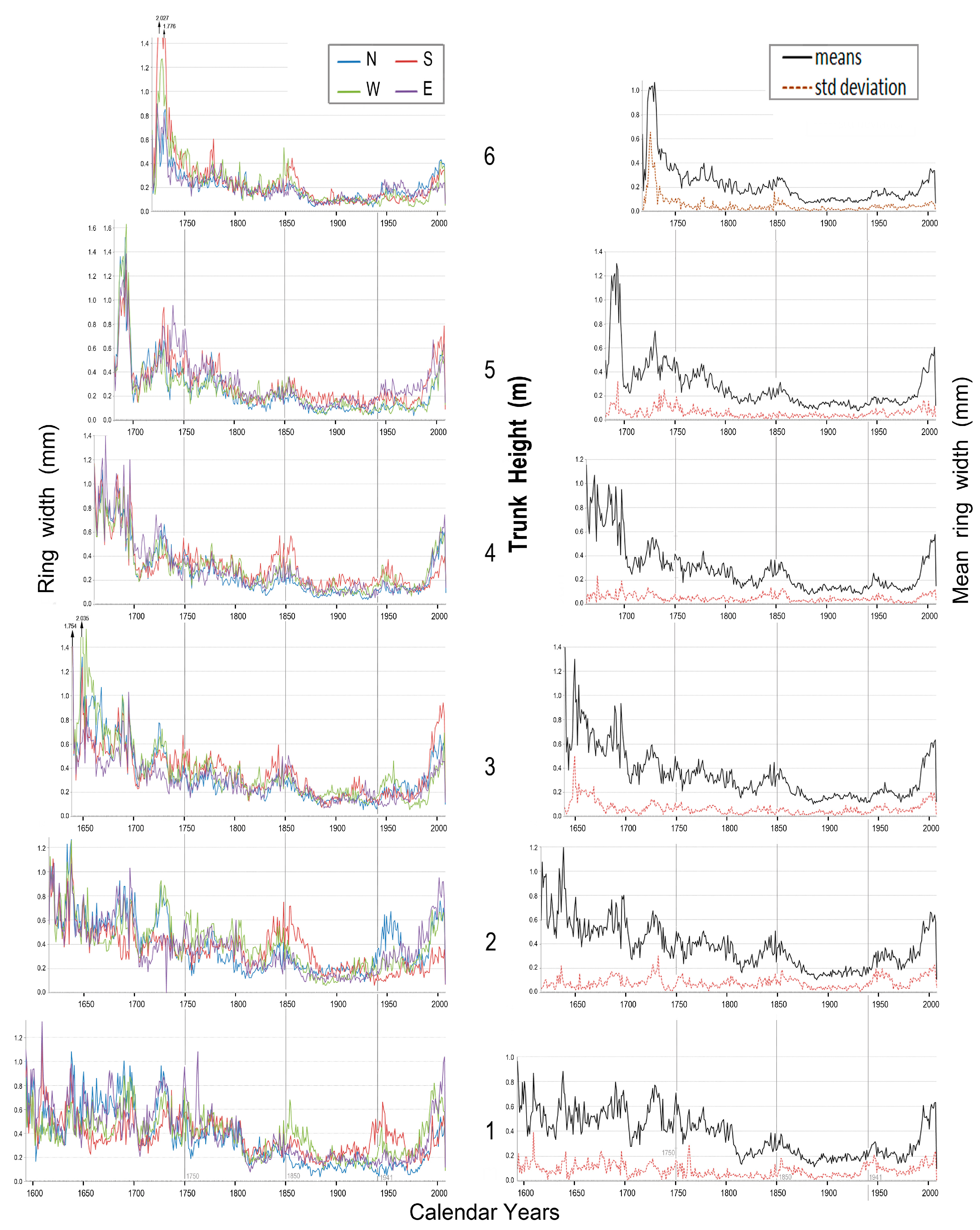
Complete sets of raw ring-width data are provided in Supplementary Materials files entitled Data1311, Data 1316, Data1320, Data 1326, Data1328 and Data1329. To exemplify the gišogenetic variation found within and between trees in this investigation, graphical data of raw ring widths are shown for tree 1311 and tree 1316, slow- and fast-growing individuals, respectively, in Figure 13, Figure 14, Figure 15, Figure 16 and Figure 17.
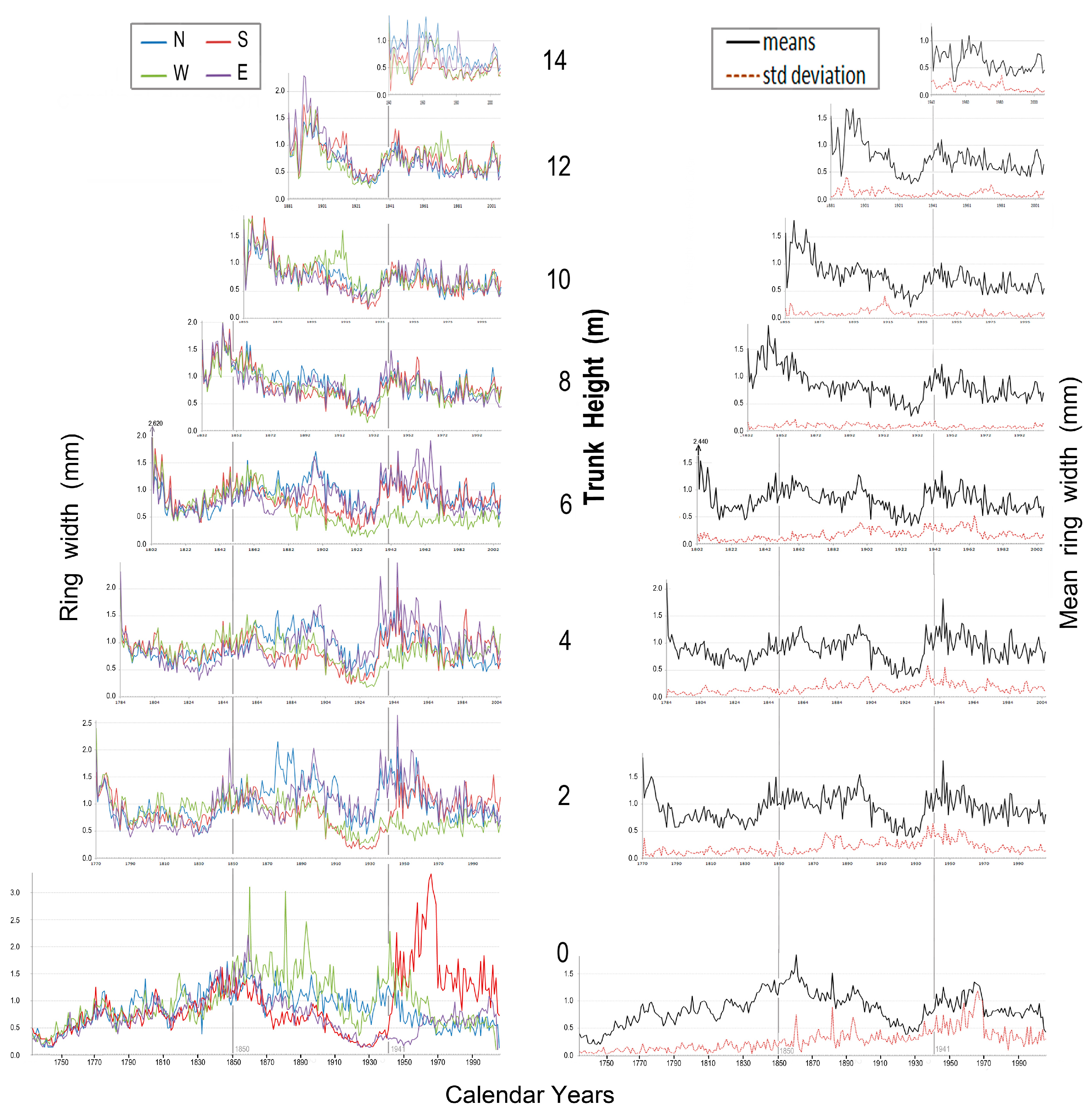
Figure 13 (left panel) presents ring-width data for each cardinal direction at heights one to six metres above ground for tree 1311. This was an 8.7 m tall tree nearly 500 years old. The right panel (Figure 13) gives mean values and standard deviations for the four sides of its trunk. After the year 1980, gišogenetic vigour became stronger throughout the trunk of this tree, but the response varied by trunk height (Figure 13).
Gišogenetic variation among the four cardinal directions occurred in some calendar-year periods. For example, during the period 1720–1740, north and east sides of the trunk experienced stronger gišogenesis at 1 m, north and west sides stronger at 2 m and 3 m, north and east sides stronger at 4 m, south and east sides stronger at 5 m, and south and west sides stronger at 6 m. Mean values and standard deviations in the right panel of Figure 12 indicate gišogenetic vigour was relatively strong in the trunk of tree 1311 over the 1600–1700 calendar period of the Little Ice Age [24,25], then declined gradually until year 1800, reached a minor peak near year 1850 and weakened again from 1875–1945.
Figure 13.
Tree 1311, left panel: ring widths of the four cardinal directions at six trunk heights; right panel: means of the four cardinal directions and their standard deviations.
Figure 13.
Tree 1311, left panel: ring widths of the four cardinal directions at six trunk heights; right panel: means of the four cardinal directions and their standard deviations.

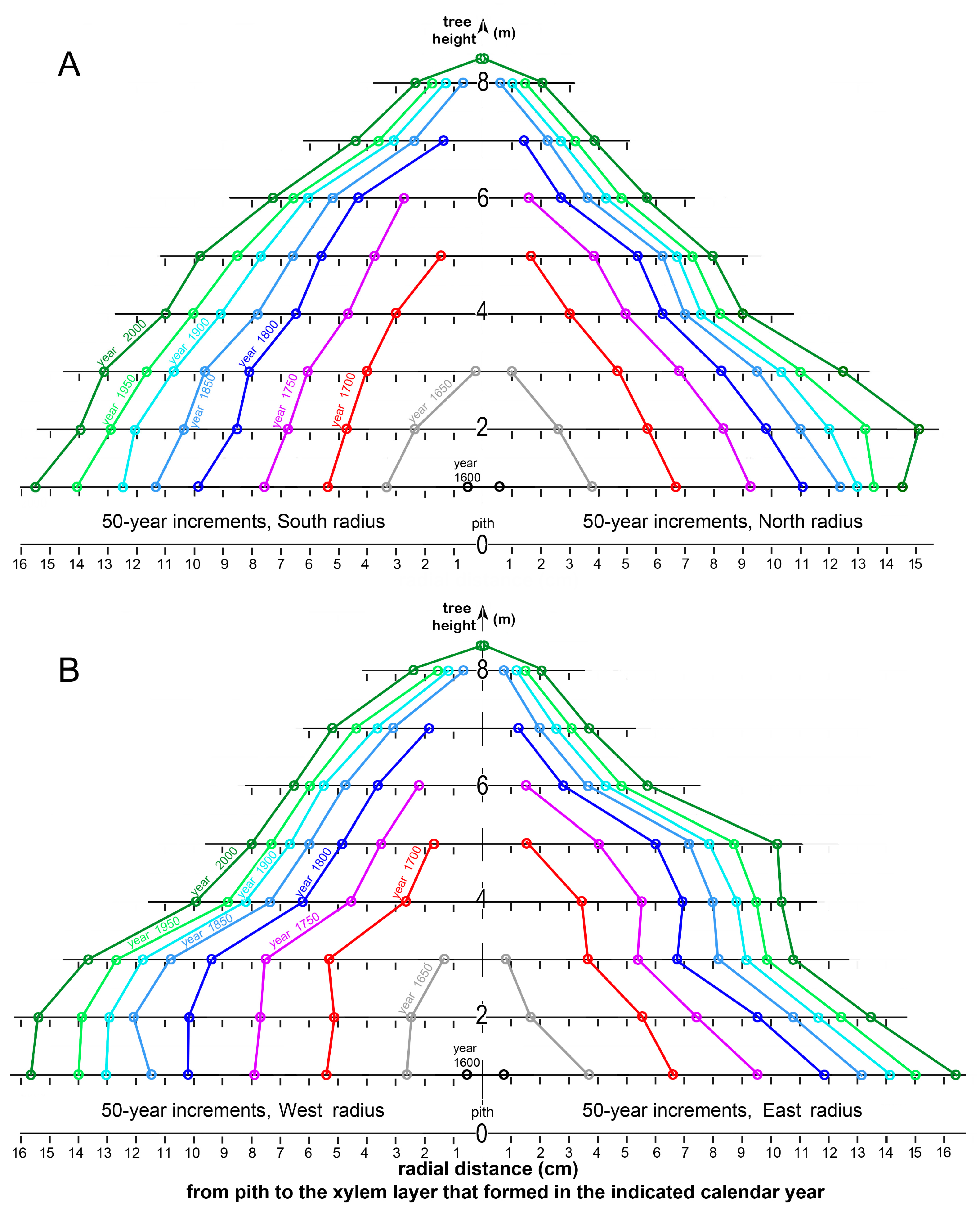
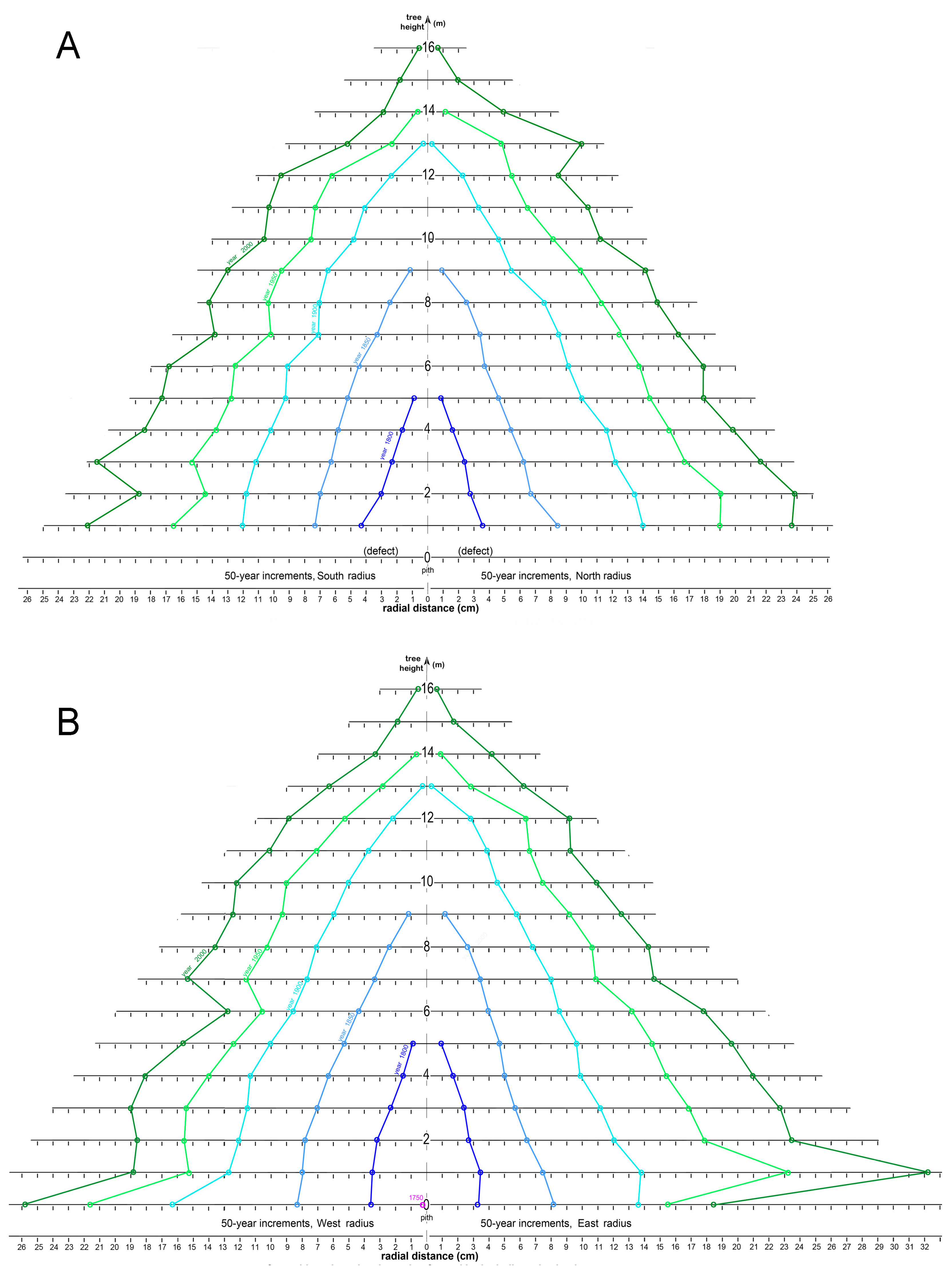
Figure 14 displays cumulative secondary-xylem increments as 50-year periods in the trunk of tree 1311 on south vs. north and west vs. east sides as the tree increased in height. Because trunk disks were sampled at 1 m intervals, trunk heights where convergence with pith occurred could not be determined with precision.
The cumulative rate of radial increment in tree 1311 was substantially greater during the period 1600–1800 than thereafter at all tree heights (Figure 13 and Figure 14). Following the 1850–1950 period of relatively slow gišogenesis, increased xylem production occurred during the 1950–2000 period, mostly after 1980 (see Figure 13).
Figure 14.
The progression of gišogenetic growth over 50-year periods on (A) the south and north sides and (B) the west and east sides of the trunk of tree 1311 as the tree increased in height.
Figure 14.
The progression of gišogenetic growth over 50-year periods on (A) the south and north sides and (B) the west and east sides of the trunk of tree 1311 as the tree increased in height.

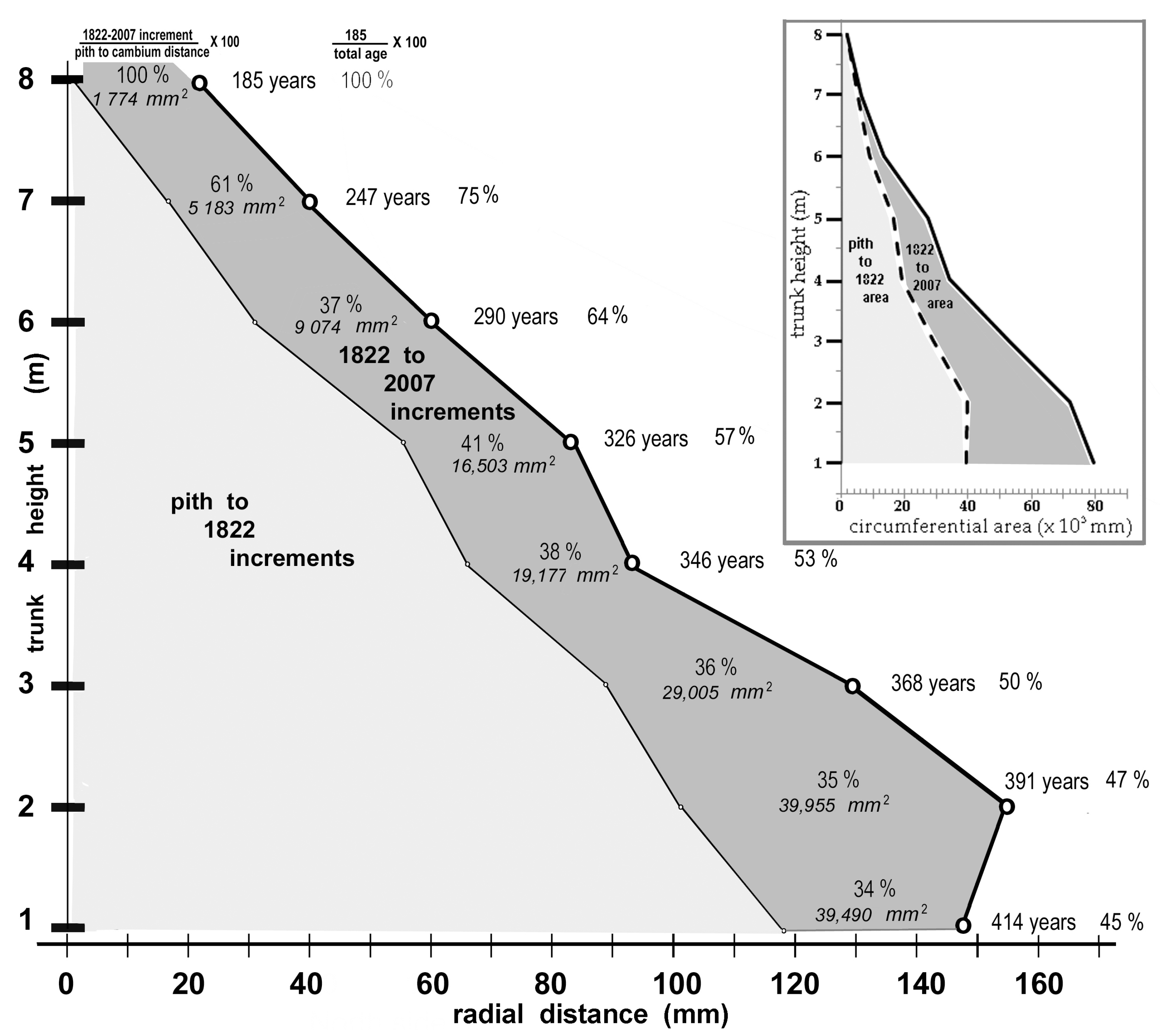
As shown in Figure 11 (inset above tree 1311), at the 8-metre trunk-height position, 0.4 m below its apex, tree 1311 contained 185 discernible secondary-xylem layers ranging from 40 µm to 280 µm in width (averages of the four cardinal directions, excluding the first two relatively wide layers bordering the pith that were 930 µm and 300 µm, respectively) in its four cardinal directions. The average radial distance for the four radii over the 1822–2007 calendar-year period was 24 mm (std. dev. 0.07 mm); therefore height growth since 1822 was limited on average to no more than 2 mm per year.
Figure 15 displays the total distance spanned by the secondary-xylem radius on the north side of the trunk of tree 1311 at each trunk height. The gišogenetic contribution to that radius assigned to the calendar-year period 1822 to 2007 is shown in dark shading. For example, at the one-metre trunk height, 229 xylem layers were deposited prior to 1822, followed by 185 layers between 1822 and 2007 (45% of 414 inferred calendar years). The more-or-less straightness of the 1822 to 2007 incremental slope can be interpreted as evidence that the 185 layers at the 8-metre height were continuous throughout the trunk. Figure 15 also gives the circumferential areas and percentage contributions to total radial expansion made at each trunk height by the 1822–2007 increments. At 8 m trunk height, the contribution to the radius was 100% but the area contribution was minor.
Figure 15.
The gišogenetic contributions in terms of radial distance and circumferential area made to secondary growth at eight trunk heights in tree 1311 during the calendar-year period 1822–2007. Area contributions were calculated by summing the radii distances of the four cardinal directions and treating the mean value as the radius of a circle.
Figure 15.
The gišogenetic contributions in terms of radial distance and circumferential area made to secondary growth at eight trunk heights in tree 1311 during the calendar-year period 1822–2007. Area contributions were calculated by summing the radii distances of the four cardinal directions and treating the mean value as the radius of a circle.

Although gišogenesis became and remained stronger in tree 1311 after the year 1980 (Figure 13), a weak to negligible increase was found in tree 1316 at all heights below 12 m (Figure 16). Moreover, in the upper crown of tree 1316, an increased gišogenetic response was delayed until after calendar year 2000.
In contrast to tree 1311 wherein gišogenesis was strongly suppressed over the calendar year period 1875–1945, tree 1316 showed a reduction only during 1910–1935. However, the two trees experienced similar reductions during 1810–1830.
The successive layers of secondary xylem that accumulated over 50-year periods on south-north and west-east sides of the trunk of tree 1316 as the tree grew in height are shown in Figure 17. Wood decay prevented resolution of all ring widths in south and north radii near ground level (0 m). Again, data are based on trunk disks sampled at 1 m intervals, and it was therefore not possible to determine with precision the height of pith convergence of the 50-year increments.
Figure 16.
Tree 1316, left panel: ring widths of the four cardinal directions at 8 trunk heights (2 m apart); right panel: means of the four cardinal directions and their standard deviations. (Note: heights shown for tree 1311 (Figure 11) are 1 m apart whereas 2 m height separations are used here).
Figure 16.
Tree 1316, left panel: ring widths of the four cardinal directions at 8 trunk heights (2 m apart); right panel: means of the four cardinal directions and their standard deviations. (Note: heights shown for tree 1311 (Figure 11) are 1 m apart whereas 2 m height separations are used here).

During the first 150 years of the life of tree 1316, trunk form was uniformly tapered on all four sides, but after calendar year 1900 irregularities began to appear (Figure 16). The rate of gišogenesis was relatively strong until 1900, slowed somewhat during 1900–1950, and began increasing during 1950–2000. The north side 9 m trunk height radial series over the calendar year period 1915–1947 in tree 1316 displayed increased growth in association with an adjoining branch. At the 2 m and 13 m heights in tree 1316, there was also accelerated gišogenesis on the north side during 1900–2000 and 1950–2000 calendar year periods, respectively, while the south sides decelerated (Figure 17A). Overall, north and east sides of the trunk were marginally more productive than south and west sides.
Figure 17.
The progression of gišogenetic growth over 50-year periods on the (A) north and south sides and (B) west and east sides of the trunk of tree 1316 as the tree increased in height.
Figure 17.
The progression of gišogenetic growth over 50-year periods on the (A) north and south sides and (B) west and east sides of the trunk of tree 1316 as the tree increased in height.

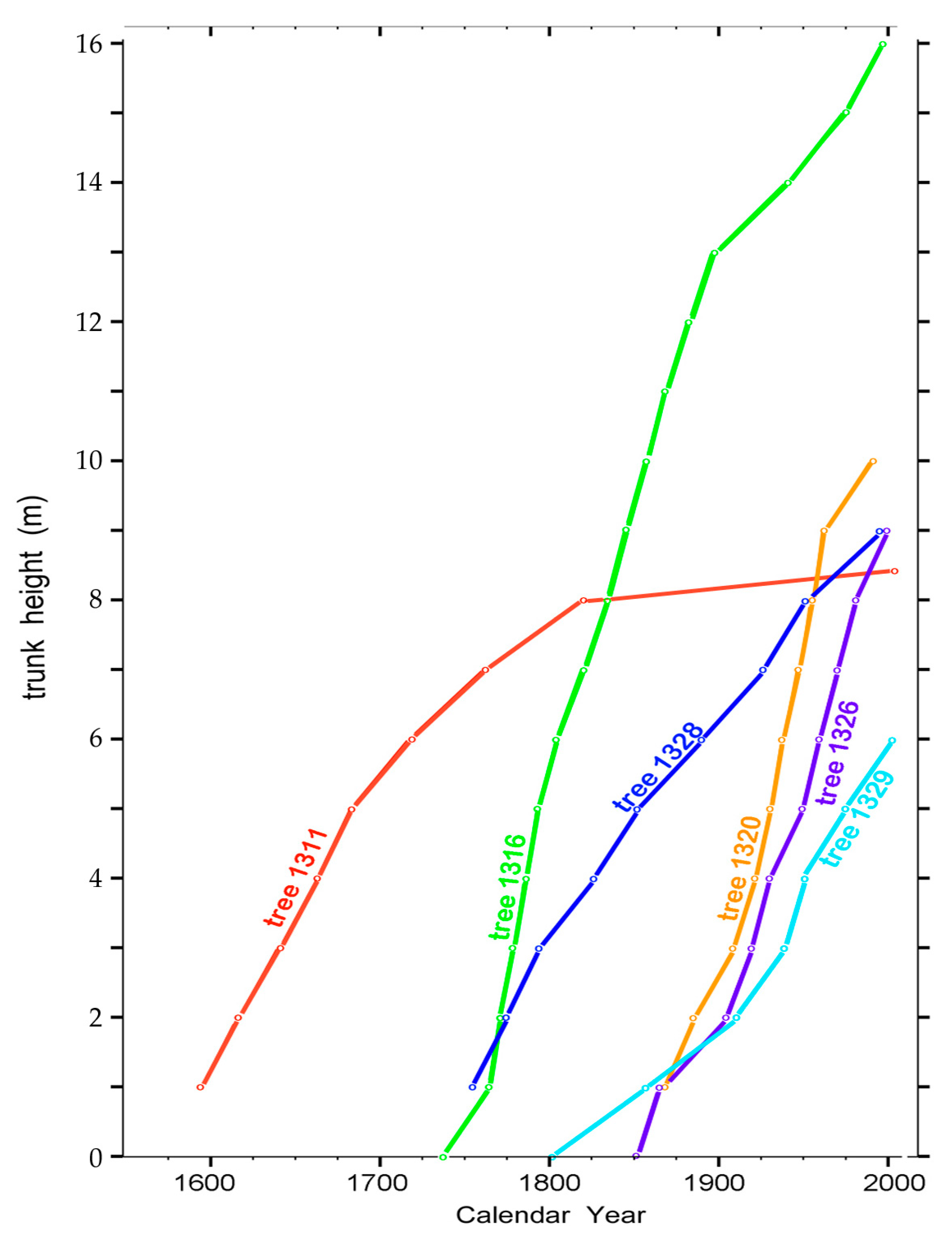
Rates of height growth of each of the six trees were deduced following calendar-year assignments to ring widths across each cardinal-direction radius. The calendar year corresponding to the secondary-xylem layer adjoining and surrounding the pith at each trunk height was considered proof that the tree had stood at least that tall in that year, enabling a plot of trunk height as a function of calendar year (Figure 18).
Figure 18.
Rates of height growth in the six investigated trees based on the year of the first xylem layer surrounding the pith at each of the indicated heights.
Figure 18.
Rates of height growth in the six investigated trees based on the year of the first xylem layer surrounding the pith at each of the indicated heights.

The height-growth data (Figure 18) indicate that the environment on the East Fork site was conducive to tree growth throughout the 1600–2000 calendar-year period. Nevertheless, as is evident by the different slopes in Figure 18, height growth rates varied among the six trees during the same calendar-year periods, indicating that each although on the same south aspect, near one another and standing unshaded, responded individually to what presumably were similar extrinsic environment conditions. Tree 1316 when a sapling was exceptional in adding, on average, more than 12 cm each year as it increased from 1 m to 5 m in height over calendar years 1761 to 1792.
Tree 1316 began its life soon after the year 1700 (Figure 18), more than a century after tree 1311. After year 1800, tree 1316 continued to grow strongly in height, whereas tree 1311 after 1800 had almost imperceptible height growth throughout the next two centuries (Figure 18). Nevertheless, gišogenesis in tree 1311 continued as in previous years (Figure 12).
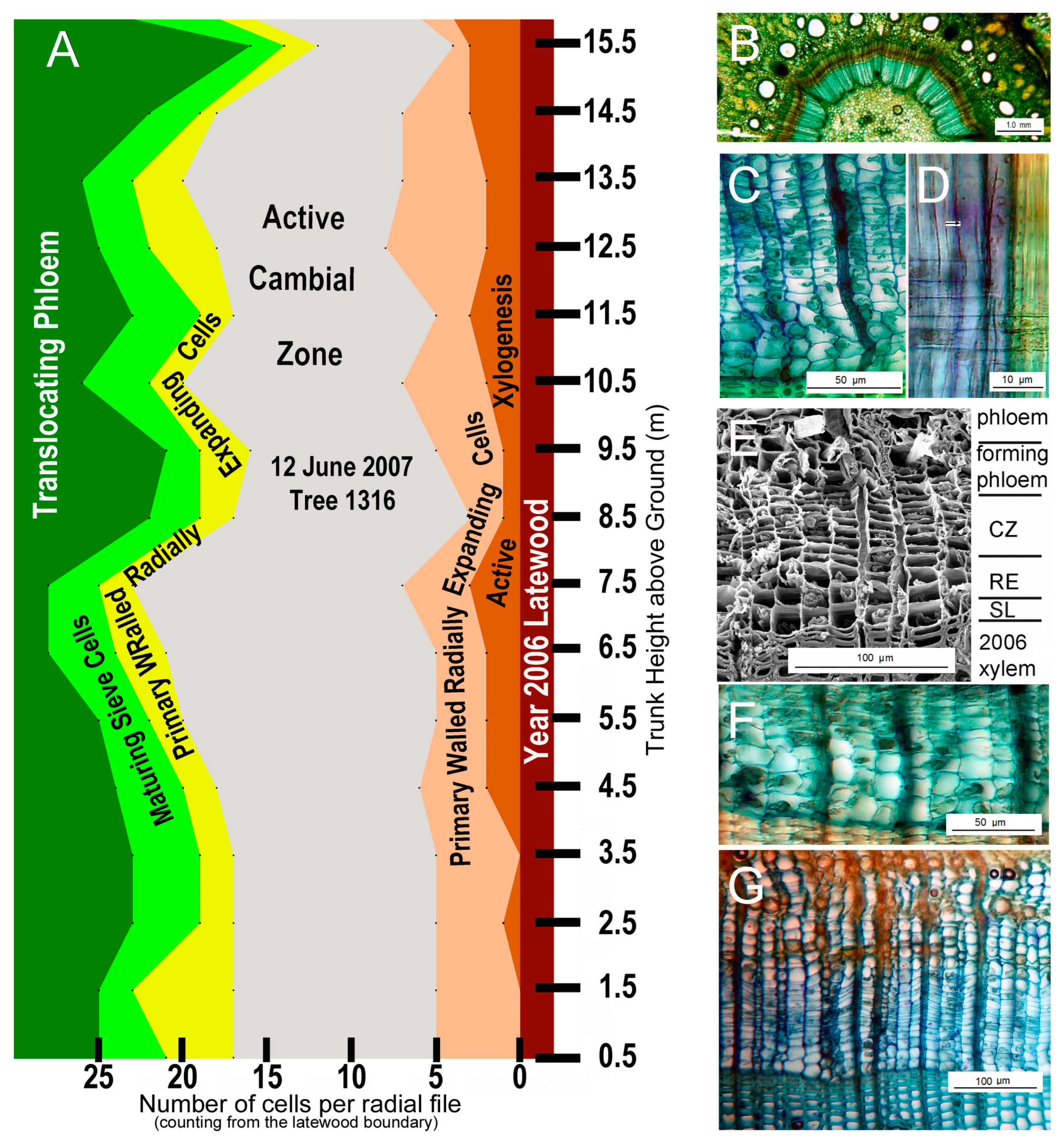
3.2. Springtime Gišogenesis Resumption in Tree 1316
The status of cambial growth, phloegenesis, gišogenesis and xylogenesis on the south side of the trunk of tree 1316 on 12 June 2007, before bud scales had separated and new shoots emerged, is shown in Figure 19. Phloegenesis was well advanced in early June (Figure 18A,B,G). Radially enlarged cambial derivatives were actively maturing into new sieve cells at all trunk positions, and additional radially expanding cells on the periphery of the cambial zone were approaching sieve-cell differentiation.
Figure 19.
The anatomical state of the cambial region at several heights above ground on the south side of the trunk in tree 1316 on 12 June 2007. (A) A graphical overview of the trunk from near ground level to tree top. (B) Transverse section showing the state in a treetop twig: the cambial zone (brown) is surrounded on its outside (centrifugally) by a relatively wide zone of new phloem cells and centripetally by a band comprising several enlarged cells per radial file becoming new earlywood tracheids through secondary-wall deposition and lignfication; the 2006 layer of mature secondary xylem (blue) is centripetal to that zone of xylogenesis. (C) Transverse section of the CZ and RE zones at 12.5 m trunk height. (D) Radial section at 12.5 m trunk height; a developing bordered pit is arrowed. (E) Scanning electron microscope image of the cambial region at 9.5 m height showing zones of developing phloem, cambial growth (CZ), xylogenesis (SL) and gišogenesis (RE + SL). (F) Transverse section of part of the CZ and all of the RE zones at 3.5 m trunk height. (G) Transverse section at 0.5 m trunk height showing active phloegenesis, cambial growth and early gišogenesis not yet attended by xylogenesis.
Figure 19.
The anatomical state of the cambial region at several heights above ground on the south side of the trunk in tree 1316 on 12 June 2007. (A) A graphical overview of the trunk from near ground level to tree top. (B) Transverse section showing the state in a treetop twig: the cambial zone (brown) is surrounded on its outside (centrifugally) by a relatively wide zone of new phloem cells and centripetally by a band comprising several enlarged cells per radial file becoming new earlywood tracheids through secondary-wall deposition and lignfication; the 2006 layer of mature secondary xylem (blue) is centripetal to that zone of xylogenesis. (C) Transverse section of the CZ and RE zones at 12.5 m trunk height. (D) Radial section at 12.5 m trunk height; a developing bordered pit is arrowed. (E) Scanning electron microscope image of the cambial region at 9.5 m height showing zones of developing phloem, cambial growth (CZ), xylogenesis (SL) and gišogenesis (RE + SL). (F) Transverse section of part of the CZ and all of the RE zones at 3.5 m trunk height. (G) Transverse section at 0.5 m trunk height showing active phloegenesis, cambial growth and early gišogenesis not yet attended by xylogenesis.

Cambial growth, phloegenesis and gišogenesis but not xylogenesis were active over the entire trunk of tree 1316 in early June (Figure 18). Xylogenesis was most advanced in treetop twigs, where a few cells per radial file were completing secondary-wall formation and lignification. At heights between 0 m and 13 m in tree 1316, only radially expanded primary-walled cambial derivatives were present. Excepting the trunk position within one metre of ground level, one or two of those cells per radial file had initiated bordered-pit formation (Figure 18D), an indication that xylogenesis was underway [23].
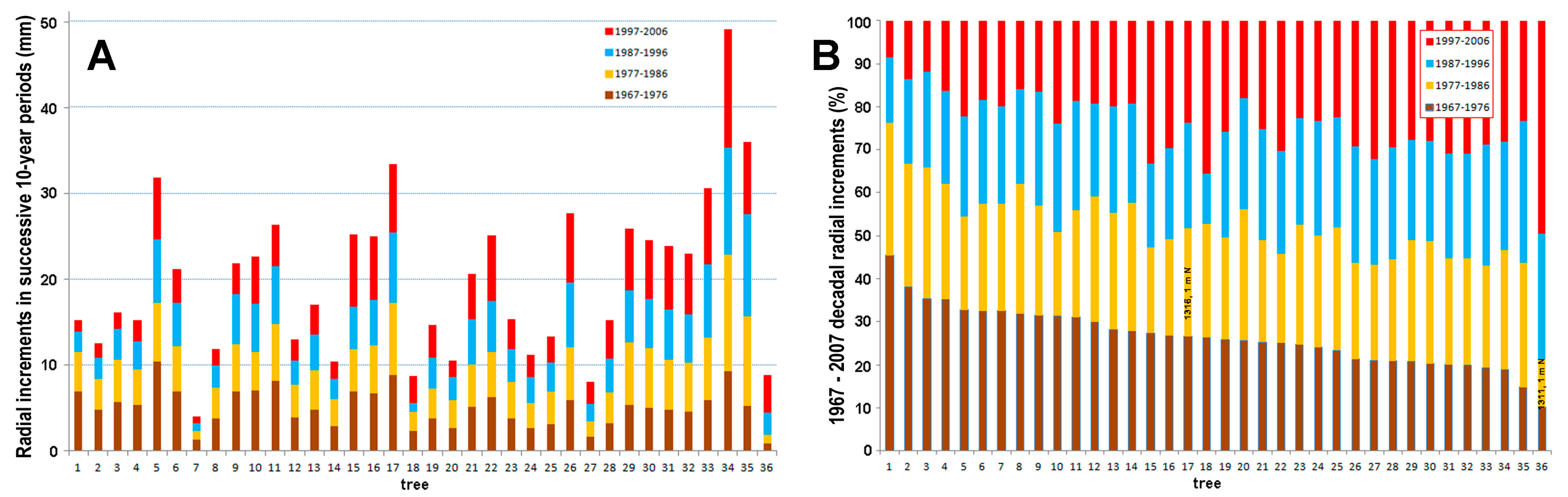
3.3. Gišogenesis during 1967–2007 Based on Core Samples from Randomly Selected Trees
Radial distances of decadal gišogenesis over a 40-year period, from the beginning of the 1967 growth layer to the end of the 2006 growth layer, are shown graphically in Figure 20A and as percentages of the total 40-year increment in Figure 20B. All 36 samples were extracted from vertical trunks of upright trees having unshaded full crowns. The major variation in gišogenetic rates is readily apparent (Figure 20A), ranging over the 40-year period from 4.0 to 49.1 mm (mean = 19.9 mm, s.d. = 9.2 mm). Of the 36 investigated cores, 13, 7, 3 and 13 had their greatest width of radial increment during the 1967–1976, 1977–1986, 1987–1996 and 1997–2006 periods, respectively (Figure 20). Unlike tree 1311, that increased in girth during those 40 years most strongly during the 1997–2006 period, many trees had their strongest gišogenesis during the 1967–1976 period (Figure 20B). The radial increments in tree 1316, towering above many others in the old-growth population, were most favoured during the 1977–1996 period.
Figure 20.
Forty-year trunk increments at 1.3 m height over the 1967–2007 calendar-year period. In addition to 34 cores, data from N cardinal directions for nominal trunk height of 1 m are shown for tree 1316 (bar 17) and tree 1311 (bar 36). Tree numbers in (A,B) apply to the same tree. (A) Raw ring-width measurements of four successive decades of gišogenesis. (B) Data of (A) converted into percentage contributions made in each decade to the total 40-year increment.
Figure 20.
Forty-year trunk increments at 1.3 m height over the 1967–2007 calendar-year period. In addition to 34 cores, data from N cardinal directions for nominal trunk height of 1 m are shown for tree 1316 (bar 17) and tree 1311 (bar 36). Tree numbers in (A,B) apply to the same tree. (A) Raw ring-width measurements of four successive decades of gišogenesis. (B) Data of (A) converted into percentage contributions made in each decade to the total 40-year increment.

Analysis of cores extracted from random trees support the interpretation that gišogenesis in tree 1311 was markedly atypical during the 1997–2006 calendar-year period. There was a relatively strong increase in gišogenetic rates in five of the six trees during 1935–1965, whereas only a weak increase occurred in tree 1311 during the same period.
3.4. Pearson’s r Values for Radial Series of Raw Ring Widths
Summarizing findings from all trees, when radial series of two cardinal directions within the same trunk disk were correlated, r values ranged from −0.08 to 1.00. Both within-tree and between-tree variation were apparent. When radial series having the same cardinal direction at different trunk heights were correlated, r values ranged from −0.44 to 0.96. When between-tree radial series were correlated, r values ranged from −0.34 to 0.88.
3.4.1. Same Tree, Same Trunk Height, Different Cardinal Direction r Values
Pearson’s correlations were done to compare complete radial series (i.e., pith to the most recently matured layer of sapwood) of the four cardinal directions at each trunk height. Table 3 and Table 4 show the results for tree 1311 and tree 1316, respectively. For example, at 4 m height in tree 1311 (Table 3), r values over the full radii varied between 0.88 and 0.94, indicating very similar gišogenetic responses on all sides. Although good correlations were found for all heights in tree 1311, tree 1316 gišogenetic responses yielded a mixture of low and high r values (Table 4).
Table 3.
Similarity of overall radial gišogenesis between cardinal directions at each trunk height in tree 1311.
Table 3.
Similarity of overall radial gišogenesis between cardinal directions at each trunk height in tree 1311.
| Tree 1311 | Correlations between Cardinal Directions | |||||
|---|---|---|---|---|---|---|
| Trunk Height (m) | Pearson’s r Value for Full Chronological Series | |||||
| N vs. S | N vs. E | N vs. W | S vs. E | S vs. W | E vs. W | |
| 1 | 0.60 | 0.79 | 0.73 | 0.69 | 0.59 | 0.73 |
| 2 | 0.62 | 0.82 | 0.79 | 0.63 | 0.73 | 0.81 |
| 3 | 0.87 | 0.86 | 0.92 | 0.86 | 0.80 | 0.82 |
| 4 | 0.88 | 0.94 | 0.95 | 0.86 | 0.89 | 0.93 |
| 5 | 0.86 | 0.84 | 0.92 | 0.89 | 0.87 | 0.85 |
| 6 | 0.81 | 0.88 | 0.80 | 0.87 | 0.89 | 0.83 |
| 7 | 0.89 | 0.90 | 0.90 | 0.91 | 0.89 | 0.87 |
| 8 | 0.79 | 0.83 | 0.87 | 0.87 | 0.87 | 0.86 |
By comparing chronological periods within series of overall high similarity, poorer correlations became apparent. For example, in tree 1311 the calendar-year period 1930–1978 was associated with changing gišogenetic rates over different cardinal directions at several trunk heights (Figure 13), and by focusing on this period r values between −0.20 and 0.52 were obtained. Numerous such low correlations were observed within the six trees, and there was no consistent calendar-year period general to all. The presence of negligible and even negative correlations at varied periods in the life of a tree is an indication that cambial derivatives on each side of the tree trunk during those periods experience differences within their intrinsic environments.
Table 4.
Similarity of overall radial gišogenesis between cardinal directions at each trunk height in tree 1316.
Table 4.
Similarity of overall radial gišogenesis between cardinal directions at each trunk height in tree 1316.
| Tree 1316 | Correlations between Cardinal Directions | |||||
|---|---|---|---|---|---|---|
| Trunk Height (m) | Pearson’s r Value for Full Chronological Series | |||||
| N vs. S | N vs. E | N vs. W | S vs. E | S vs. W | E vs. W | |
| 1 | 0.66 | 0.44 | 0.61 | 0.56 | 0.50 | −0.08 |
| 2 | 0.36 | 0.69 | 0.34 | 0.48 | 0.52 | 0.18 |
| 3 | 0.56 | 0.66 | 0.56 | 0.68 | 0.33 | 0.13 |
| 4 | 0.50 | 0.66 | 0.49 | 0.60 | 0.27 | 0.43 |
| 5 | 0.55 | 0.69 | 0.57 | 0.61 | 0.53 | 0.31 |
| 6 | 0.62 | 0.73 | 0.43 | 0.65 | 0.71 | 0.26 |
| 7 | 0.76 | 0.83 | 0.78 | 0.85 | 0.81 | 0.76 |
| 8 | 0.88 | 0.86 | 0.89 | 0.86 | 0.92 | 0.85 |
| 9 | 0.38 | 0.48 | 0.41 | 0.91 | 0.88 | 0.92 |
| 10 | 0.90 | 0.85 | 0.88 | 0.90 | 0.80 | 0.75 |
| 11 | 0.91 | 0.83 | 0.92 | 0.83 | 0.95 | 0.85 |
| 12 | 0.93 | 0.90 | 0.81 | 0.86 | 0.78 | 0.72 |
| 13 | 0.37 | 0.56 | 0.27 | 0.59 | 0.63 | 0.45 |
| 14 | 0.74 | 0.79 | 0.79 | 0.62 | 0.73 | 0.67 |
| 15 | 0.86 | 0.82 | 0.89 | 0.88 | 0.82 | 0.84 |
| 16 | 0.85 | 0.60 | 0.91 | 0.77 | 0.82 | 0.40 |
3.4.2. Same Tree, Same Cardinal Direction, Different Trunk Heights: r Values
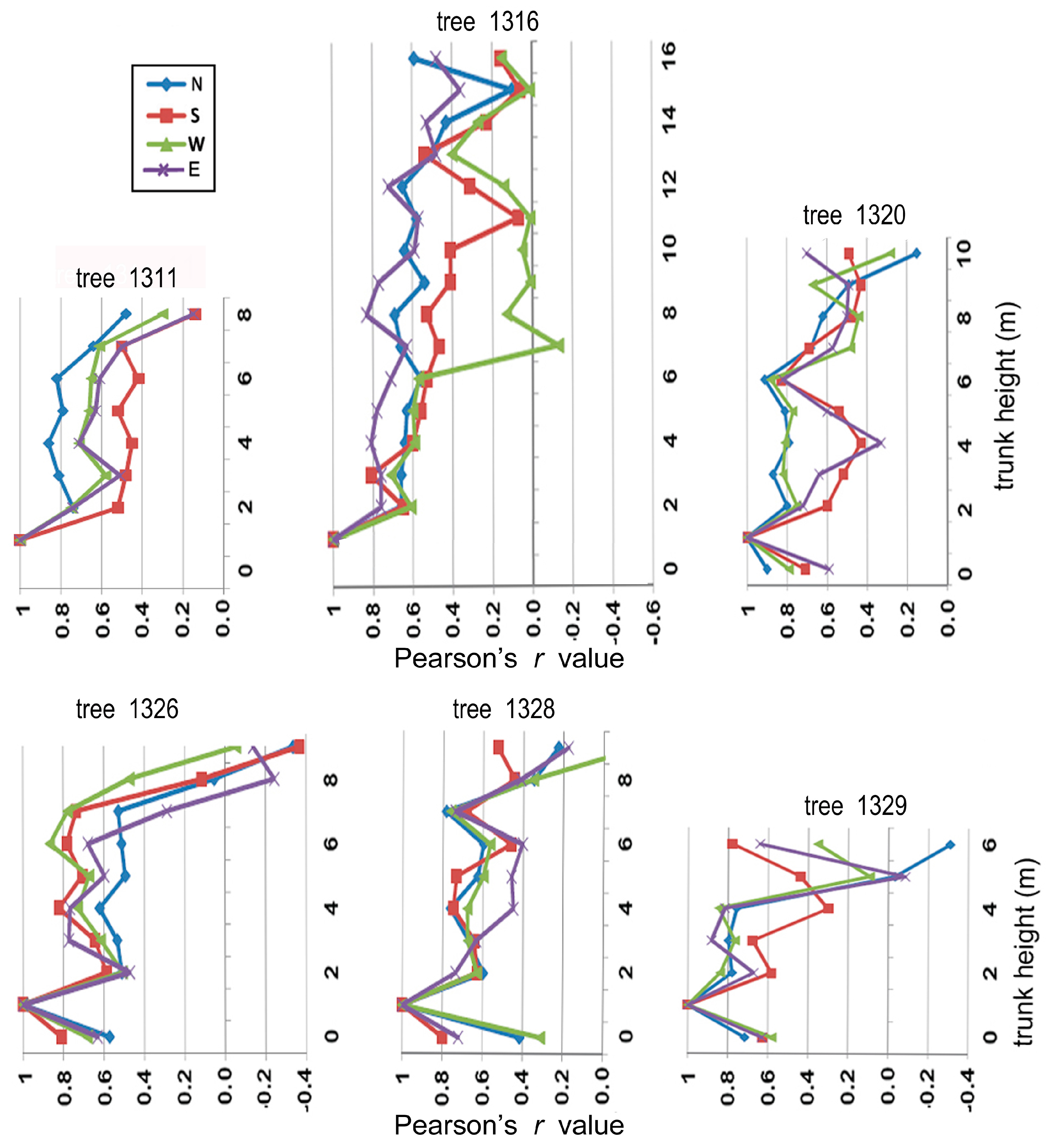
Figure 21 displays examples of within-tree variation in Pearson’s r values found for all six trees, after correlating radial series at other trunk heights with those at the 1 m nominal trunk-height position. The correlations showed a tendency to decline acropetally (Figure 21), and the lowest r values were often generated after comparing uppermost disks with disks near ground level.
The gišogenetic rates on the west side of tree 1316 were poorly if at all correlated with the 1 m trunk height and clearly deviated from the correlations found for the other three sides at heights above 6 m (Figure 21). In tree 1320, the gišogenetic-rate correlations of the south and east sides became distant from those of north and west sides at trunk heights between 2 and 5 m. The four cardinal directions yielded similar r values for trees 1311, 1326 and 1328 at all trunk heights, whereas r values for the south and north sides deviated from those of east and west sides in the treetop of tree 1329.
Figure 21.
Variation in Pearson’s r values produced by correlating radial series at 1 m trunk height with those of corresponding chronology and the same cardinal direction at other trunk heights.
Figure 21.
Variation in Pearson’s r values produced by correlating radial series at 1 m trunk height with those of corresponding chronology and the same cardinal direction at other trunk heights.

3.4.3. Between-Tree Comparisons
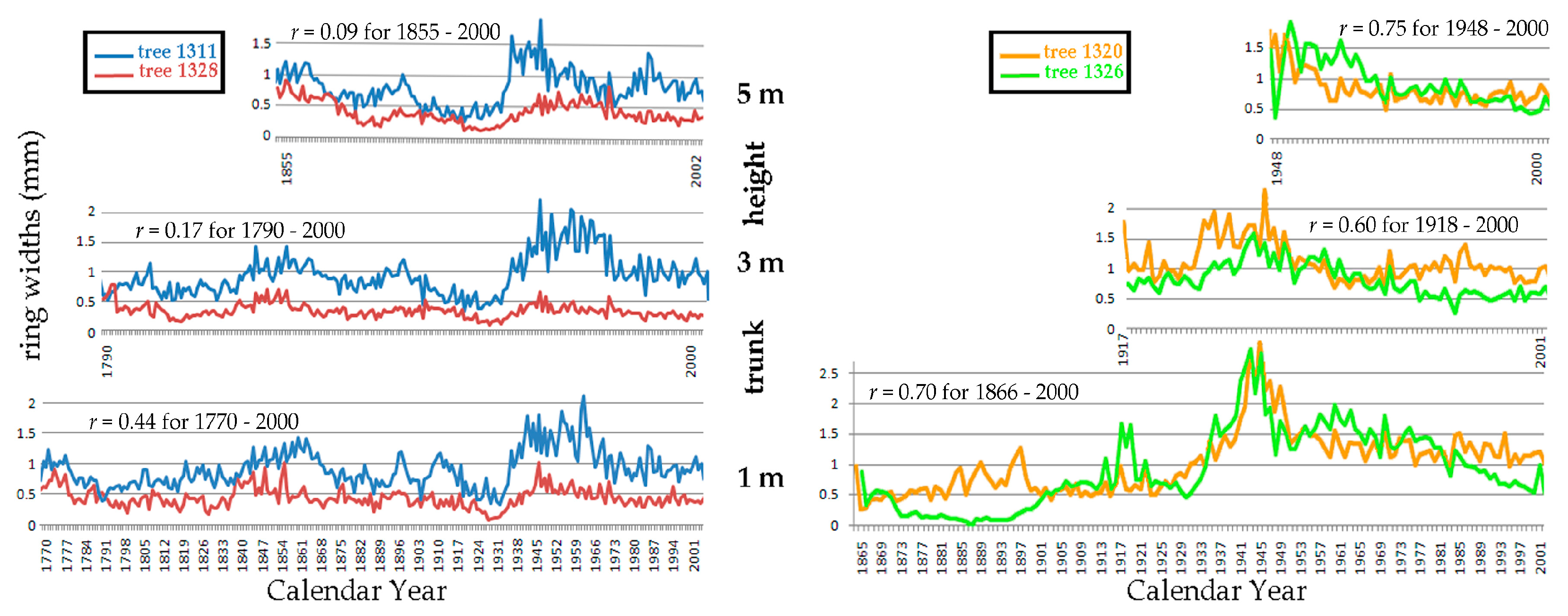
Correlations between trees of chronologically corresponding radial series at the same trunk heights yielded Pearson’s r values ranging from −0.34 to 0.88 for all six trees. Figure 22 provides examples of six such correlations. Low r values are shown in the left panel and stronger correlations in the right panel.
Figure 22.
Between-tree correlations of gišogenetic vigour in chronologically corresponding radial series on the south side of each trunk at three trunk heights. The left panel compares tree 1311 with tree 1328, and the right panel compares tree 1320 with tree 1326.
Figure 22.
Between-tree correlations of gišogenetic vigour in chronologically corresponding radial series on the south side of each trunk at three trunk heights. The left panel compares tree 1311 with tree 1328, and the right panel compares tree 1320 with tree 1326.

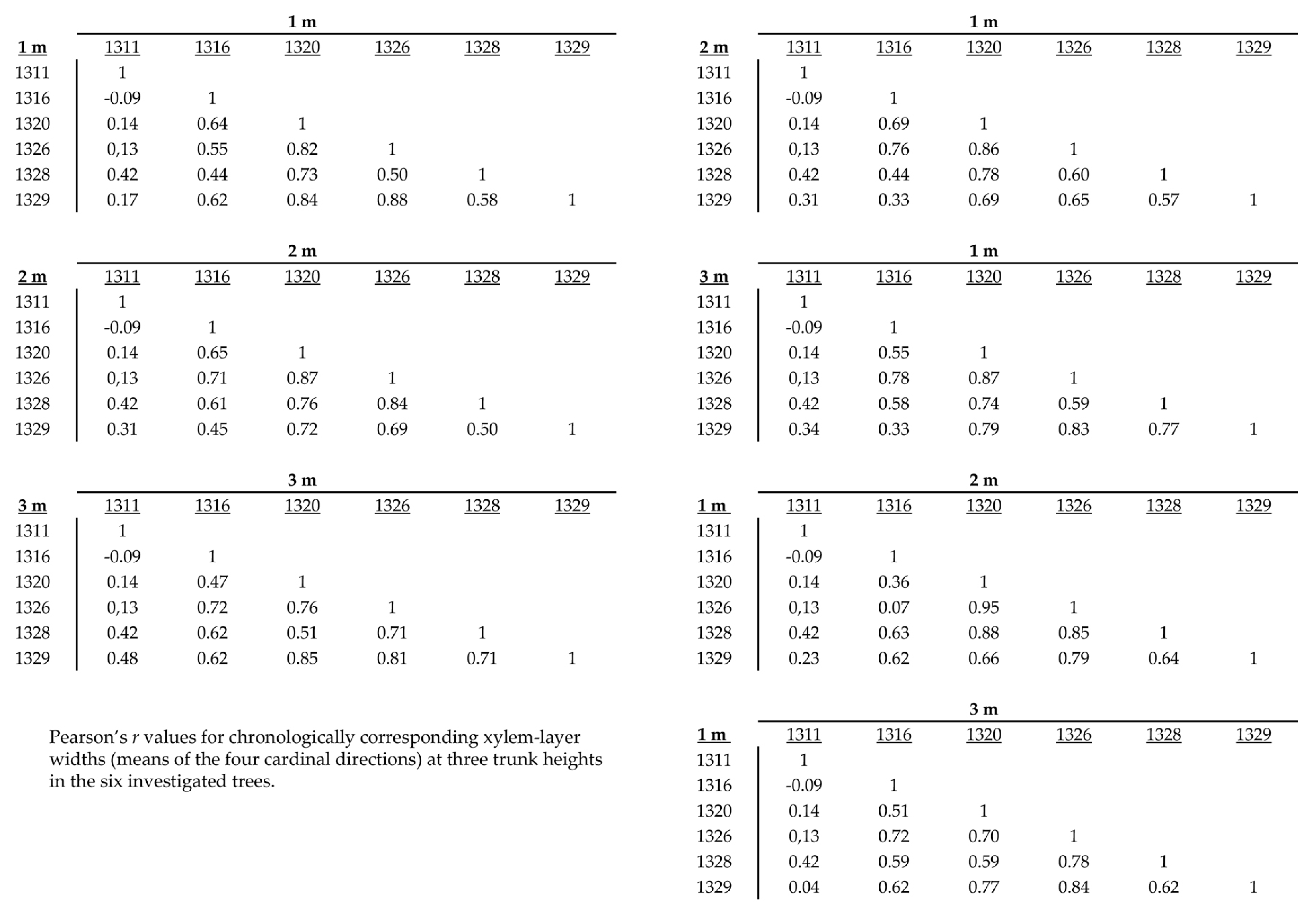
Figure 23 summarizes correlations found between trees by comparing trunk heights of 1, 2 and 3 m. Pearson’s r values ranged from 0.95 to −0.09 (mean 0.53, std. dev. 0.28) for the 105 comparisons of chronological ring-width series shown in Figure 23. Tree 1311 series were in least agreement with any of the other five trees, but those of tree 1316 also resulted in relatively weak correlations.
Figure 23.
Summary of between tree gišogenesis (ring-width) correlations for all 6 trees, showing only trunk heights 1, 2 and 3 m.
Figure 23.
Summary of between tree gišogenesis (ring-width) correlations for all 6 trees, showing only trunk heights 1, 2 and 3 m.

4. Discussion
This is the first description of white-spruce gišogenesis in this remote region near the southeastern tip of Eastern Beringia. To our knowledge, it is also the first Yukon study to examine gišogenesis systematically at successive trunk heights, from near the ground level to tree top, over the four cardinal directions at each height. Trunks of each of the six trees were examined carefully, and the concordance in number of xylem layers resolved over the four cardinal directions at each trunk height enabled confidence in the existence of completely circumferential layers, hence the inference of chronological correspondence. This four-sided sequential approach minimized the common difficulty associated with analysis of cores, where within-tree variation may appear to exist because some trees have missing ‘rings’ at the examined trunk height. Considering the very short growing season on site, production of a general late-season secondary annual layer of xylem through lammas growth was considered improbable [26].
Numerous earlier investigations have examined wood in white-spruce trunk cores and root collars in efforts to deduce past climate of northwestern North America. That was not the aim of this investigation, and therefore transformations such as “application of detrending techniques … to remove tree age related (biological) trends from width or density data” were not done [27,28]. All aspects of diameter growth in trees remain imperfectly understood in terms of their regulation by both extrinsic and intrinsic factors and, therefore, only raw ring-width data were analysed by us.
There is earlier evidence for variation in ring-width correlations between sites [29,30]. For example, 83% of northern Alaska white-spruce trees contradict the uniformitarian ‘principle,’ a dendroclimatological concept that the relationship between a region’s climate and tree growth should be linear and stable over time for the general population [28]. More recently, variation between two southwestern Yukon mountain ranges less than 100 km apart was reported [29].
Evidence for gišogenetic variation within localized populations has also been reported, again based on ring-width data from cores extracted at one trunk height [30,31]. Older trees evidently yield a different gišogenetic response to that in younger trees on the same site, perhaps explainable in terms of physiological change at the sampled height associated with aging [30].
The six trees we investigated were randomly chosen, and they began their lives over a span of 300 years, from sometime in the 1500’s to the mid-1800’s (Figure 18). Several correlations revealed higher gišogenetic similarities between younger trees than did correlations between older and younger trees, although the largest tree yielded a range of Pearson’s r values from low to high in that respect (Figure 23). Thus, our investigations further confirm that gišogenesis can vary substantially between trees within the same calendar-year period, despite all residing at one location and experiencing similar extrinsic environment. The observed within-tree reductions in the strength of Pearson’s correlation between older trunk positions, such as at breast height, and younger positions nearer the treetop, may also be related to this age factor.
Although no climatic data exist for the investigated site during the 1967–1976 calendar-year period, there is certainty that both an East Fork valley glacier and snow on north slopes persisted throughout the summer months, that the willow and birch shrubs were greatly compromised in their abilities to grow, and that all of those features subsequently changed in support of a flourishing plant community. However, despite the evidence for on-site warming sometime after 1976, the ring-width data from cores revealed that many trees had stronger gišogenetic vigour during the 1967–1976 period than thereafter, although another significant fraction of cores yielded stronger gišogenetic vigour in decades after 1976. Again, variation in annual diameter-growth rate appears to be the rule rather than the exception on this East Fork site.
Based on the oldest investigated tree, gišogenetic vigour during the 1600–1700 period of the Little Ice Age remained relatively strong. However, after 1850 when that globally cold period was fading [24,25], vigour declined. This was surprising and indicates the need for investigation of additional trees on this site.
Water is essential if cambial growth is to occur [32]. Despite the semi-arid precipitation inputs and freezing night-time air temperatures affecting the East Fork site during the growing season, thawing ground ice can be a vital source of water [33]. Our temperature data at different soil depths indicate that liquid water is probably available to root systems even during winter. However, it remains near the freezing point even during summer, and there can be negative effects on tree growth when soil water remains perennially cold [34]. The magnitude of such negative effects evidently varies the adaptive tolerance of each genotype [32,34].
All observations of the variation in white spruce gišogenesis on the East Fork site point to the existence of considerable genetic diversity. Each tree’s genome evidently is in possession of unique attributes of physiological adaptation endowing its intrinsic gišogenetic vigour. Similar observations apply to survival fitness in this region [35]. There presently is no other physiologically well-substantiated explanation for individualistic growth responses within the same climatic region being the norm.
A population localized to a particular geographic realm adapts, through natural selection, to survive in that particular environment [35]. It has been well established from provenance tests in many locations (although evidently not yet in Yukon [36]) that white-spruce trees of different geographic origin when grown side by side on the same site vary in their abilities to survive and grow [37,38]. This variation is generally explained in terms of differences in geographic environments and degrees of adaptation [38,39,40].
Several Yukon dendroclimatology investigations into secondary xylem of white spruce have been done in the Klondike region to the north and in the Shakwak Valley region to the south of the East Fork site. However, neither of those locations holds old-growth populations comparable to the East Fork site. The old growth of the Klondike was harvested during the gold-rush period at the beginning of the 20th century, such that most trees presently there date back to the early 20th century and probably none earlier than 1850 [41]. The white-spruce populations in the Shakwak Valley have also been affected by harvesting, forest fires and insect epidemics; moreover, the climate of that region is not as severe as that of the southern Dawson Range holding the East Fork site [14]. The population on the East Fork site is essentially an isolated island of white-spruce trees near the altitudinal limit of tree survival. Pockets of similar old growth not yet accessible by road have been noted in other remote upland locations within the Dawson Range [13]. Considering the extreme environment affecting this region and the fact that these pockets consist only of spruce trees, it can be suggested that the trees are exceptional survivors having high probability of being descendents of persistent local refugia [9,12,35,42]. If they are not derived from ancestors that stood on the same site, they could be from seeds transported from other refugial locations within Eastern Beringia by wind or by restricted-locale birds such as the boreal chickadee [43]. Considering the thousands of years that have passed since the LGM, it seems probable that others within populations such as the East Fork site will have originated from post-glacial re-entry into Yukon [44]. In other words, the observed variation in gišogenetic responses may be linked to different geographic ancestries and phenological adaptations to distinct climates.
Cambial growth and gišogenesis have frequently been linked on the assumptions that cambial growth must occur annually in a living tree and that wherever it occurs gišogenesis must follow. However, there is no proof that vascular cambium in order to persist as a living meristem must reactivate each year. Based on current physiological understanding, it can be suggested that if a tree is in possession of both translocating phloem and conducting sapwood, both of those tissues being in a functional state, its vascular cambium although dormant could sustain itself (through phloem unloading and a continuing supply of sapwood solution) such that occurrence of gišogenesis during each successive growing season were unessential.
If cambium in evergreen trees in fact is competent to endure multi-year dormancy during extended periods of spring/summer low temperatures, variation in the cambium’s ability to reactivate at all positions or throughout the trunk during the growing season could also vary among ancestral lines. In this vein, between-tree variation on the East Fork site could have its explanation in terms of gišogenesis occurring only in the years when conditions were attuned for a particular lineage. It is difficult to identify a physiological reason to dismiss such speculation as invalid.
Production of cambial derivatives is the necessary prelude for gišogenesis, but production of derivatives on the interface of the cambial zone with the mature functional phloem in support of phloegenesis has phenological priority [45]. That was evident by analysis of tree 1316 soon after cambial growth had commenced. Numerous experiments with bark-ringed trees have revealed their capability to survive, for a year or more, after being girdled. For example, if a spruce trunk is girdled below its live crown, the tree does not die instantly or even quickly, because the supply of water to the needles is not interrupted [46]. However, when the phloem translocatory pathway is interrupted, delivery of photosynthate to the root system is prevented; it must draw on its storage reserves and although cambial growth is still possible in the roots the nature of gišogenesis becomes altered [46]. Unless the girdled tree is root grafted to another, the phloem blockage eventually results in death of the root system followed by the whole tree.
Based on the foregoing, if no gišogenesis were possible because of cold dry environmental conditions persisting throughout the growing season on the East Fork site during some past periods, for example, during the 1967–1976 period when such conditions were plainly evident, ring widths would be ‘silent’ rather than merely narrowed within chronological series. Plausibly, multi-year protracted cambium dormancy, or centrifugal-only production of cambial derivatives in support of phloegenesis, explains the variation in gišogenesis among the six investigated white-spruce trees. If silent, calendar-year assignments based on counting xylem layers backward toward the pith would be incorrect and, if only some trees experienced periods of non-gišogenesis, between-tree variation in chronological series of ring widths would be increased.
The pith-to-cambium radius in the 8-m disk of tree 1311 (Figure 11, inset) was 22 mm on north and east sides and 25 mm on south and west sides, and yet under the microscope it contained 185 distinguishable layers of xylem spanning calendar years 1822 to 2007. The stem at 8-m height was near green leaves, and gišogenesis for its occurrence depends on contributions from leaves, i.e., both products of photosynthesis and regulatory factors [47]. If those essential products exported from the leafy crown varied, such that under limiting extrinsic conditions gišogenesis were possible only in the upper region of the tree containing the greatest density of leaves, this could be yet another explanation for variation between trees and perhaps also within a tree. However, the data from tree 1311 indicate just the opposite, as the gišogenetic increments display a more or less uniform slope over the entire trunk from 1822 to 2007 (Figure 15).
As a trunk increases in diameter, the area and number of cambial cells concomitantly increases, hence requiring the supply of more nutriment, and conceivably also more regulatory factors, to achieve a gišogenetic increment equivalent to that produced higher in the smaller diameter trunk. In fact, the radial width of the 1822–2007 increment was greater in the lower than in the upper trunk region (Figure 15). Moreover, when tree 1311 was felled, living needle-bearing branches were attached to its trunk near ground level (Figure 11). Thus, a limitation of photosynthate or regulatory factors appears to be an improbable explanation for the poor correlations in gišogenetic rates between tree 1311 and the other five trees.
One additional genotype-focused explanation for between-tree gišogenetic variation is also relevant to within-tree gišogenetic variation. Invariance, or constancy, of the somatic genome inherited by the seed embryo during vegetative development has been a dogma of the biological sciences over the last century. Thus, the cambium within each tree has been assumed to be a singularly uniform somatic genotype. However, a remarkable amount of genomic variation, perceivedly arising through mutation, has recently been described within old-growth Sitka-spruce trees [48]. Acquisition of somatic mutations evidently is an environmentally induced random process, cumulative with passage of years, and distinct genotypes at different tree heights could modify gišogenetic vigour. How somatic variation may modify the nature of cambial growth and gišogenesis remains to be clarified.
The extrinsic environment affecting trees is relatively easily measured, but in order for it to affect secondary growth it must be transduced directly through the periderm, or indirectly via the translocatory and transpirational streams, into the cambial region. In other words, the intrinsic environment is immensely more complex than the extrinsic environment affecting a tree trunk. Moreover, at the level of cell biology, the intrinsic environment is continually in motion within every living cell, as well as between cells and over long distances.
The precise explanation(s) for gišogenetic variation at the East Fork site is presently uncertain, needing of additional research. The observed variation between cardinal dictions at the same trunk height and between different trunk heights can nevertheless be hypothetically rationalized in physiological terms. A brief review is needed to explain this.
In the late 1800’s, the favored hypothesis for explaining how trees make wood was that dormant cambium became activated in springtime by an intrinsic factor called “Wuchsstoff” (German for ‘growth substance’) that was produced after budbreak by extending shoots and developing leaves [49,50,51,52]. Wuchsstoff became known in the early 20th century as ‘auxin’ (a Greek word meaning ‘to grow’) [52]. Indol-3-ylacetic acid (IAA) was the first auxin to be extracted and purified from plant tissues [52,53,54], and IAA subsequently became synonymous with auxin in a great many research publications. Several other plant metabolites having activity similar to that of IAA in auxin bioassays have been discovered [52,53,54] but, excepting indol-3-ylbutyric acid and the aspartic-acid conjugate of IAA [55], it remains uncertain if any endogenous to cambium can have the effects of exogenous IAA on either xylogenesis or gišogenesis.
The Wuchsstoff hypothesis included the concept that secondary growth always followed and was dependent on primary growth. In the 20th century it evolved into the concept that an apical supply of IAA exported from extending shoots and developing leaves was essential for induction of cambial growth and gišogenesis, and that each spring the reactivation process propagated basipetally through twigs into the trunk and on into roots [56]. This became the auxin theory for cambial growth. However, abundant evidence has shown it to be an improbable theory at the very least in need of major revision.
Cambial reactivation in conifers older than a few years was observed to occur first in the trunk near the base of the live crown, rather than at bud bases, whereas xylogenesis occurred first in the tree top [56,57]. The same reactivation sequence was evident in this investigation. Evidence was found for continuing IAA presence and basipetal movement during tree dormancy, showing that primary growth need not occur in order for IAA to be provided the cambium [58]. Moreover, after warming localized regions of stems of dormant trees standing in a chilled environment, cambial reactivation and xylem production were found to occur at the warmed site, while buds and cambium elsewhere remained dormant [55], an observation that has now been confirmed in various species [59,60,61,62].
Yet another difficulty with the auxin theory emerged with the observation that although exogenous auxin was effective in promoting some gišogenesis when applied to stem segments two years of age and younger, neither cambial growth nor gišogenesis occurred in older stem segments [47,55,63]. In older cambia, a combination of auxin, gibberellin and cytokinin stimulated minor cambial growth; however, neither that triplet combination nor any pair of them promoted xylogenesis [63]. In vitro investigations with one-year-old living but dormant stem segments lacking apical meristems and leaves, after having their apical ends infused with a sterile solution containing auxin and/or other phytohormones, together with perceivedly ample inorganic and organic substrates and ideal environmental conditions for cambial growth, underwent gišogenesis for only a limited period compared to the ongoing activity normal to similar sized stems in situ on whole plants [47,55,64]. Similarly, although limited success in promoting in vitro gišogenesis in older cambia was achieved, sub-culturing was ineffective in sustaining cambial growth [65]. Moreover, it was discovered that plant cells could differentiate directly into tracheary elements independently of preceding cell-division activity [22,47,66].
Numerous transmissible organic compounds when provided exogenously under controlled experimental conditions to stem segments have been observed to have effects on gišogenesis. Of those, ethylene has long been recognized to modulate IAA synthesis, transport and signaling [67,68]. There is increasing evidence that ethylene rather than IAA actually initiates xylogenesis [69]. Cambium two years of age and younger was observed better able to generate ethylene, and seven-year-old cambium was only competent to produce ethylene from its precursor (1-aminocyclopropane-1-carboxylic acid) if auxin was also present [67].
Gibberellins [63,70], cytokinins [63,71], abscisic acid [72], brassinolide [73], strigolactone [74], jasmonic acid [75], flavonoids [76], cis-cinnamic acid [77], salicylic acid [78], coumarins [79], dihydroconiferyl alcohol [80] and dehydrodiconiferyl alcohol glucoside [81] have all been identified as endogenous regulators. Plasma membrane and extracellular arabinogalactan proteins [82,83], various peptides [84] and additional physical and chemical agents have also been implicated [3,47,54,55,85]. In view of the number, their diverse effects, interactions and the possibility that each may vary independently in both abundance and phenology, gišogenesis regulation clearly is highly complex. It could be argued that variable expressions within and between individual perennial woody plants should be expected as the norm, and this is amply evident in the diverse woods in existence [2]. In this light, it becomes challenging to explain how strong correlations have sometimes been found within a geographic region. All of these endogenous regulators are awaiting more intensive investigation.
It is well established that traumatic longitudinal resin canals in the Pinaceae are caused by wounding, insectivory, etc., [86,87]. Longitudinal resin canals were rarely seen in the disks of the investigated trees, and bands of them indicative of trauma were seen only where former scarring of the bark had occurred. On the basis of the six trees investigated, it can be suggested that the population either has resistance or is sufficiently isolated to have been shielded from exposure to insects such as bark beetles. However, it was difficult to find good seeds in this population as cone insects are abundantly present.
In the investigated East Fork population, frost cracks were rarely noted, a surprising observation as longitudinally split trunks are common at lower Yukon elevations. Frost cracks are separations parallel to the grain in the radial plane of trunks arising from tension stress perpendicular to that plane under freezing conditions [88]. The presence or absence of frost cracks in a population is diagnostically informative of the variable strength properties of wood. With additional research, this population could enable greater insight into how to better manage those properties.
5. Conclusions
Pearson’s correlation data at successive trunk heights in six trees have indicated, as a general conclusion, that rates of gišogenesis can vary substantially both within and among genotypes of the East Fork population, despite the extrinsic environment of the site being overtly uniform. This variation was further substantiated by examination of a number of trunk cores extracted at breast height from other trees on site. It is apparent, therefore, that dendroclimatological inferences based on sampling of old-growth white spruce on this East Fork site would have high uncertainty. It would be of interest to discover if similar variation exists in the other pockets of old-growth white spruce scattered across the mountainous Dawson Range of Yukon Beringia.
The variation in gišogenetic rate correlations between the four cardinal directions and between different trunk heights within individual trees ranged from strong to no correlation. The existence of such a range allows the conclusion that during periods when the correlation is poor, cambial growth at each trunk location can respond independently of other locations. It follows that the intrinsic physiological factors regulating gišogenesis must vary from one trunk position to another within the same growing season. Research is needed to identify those factors and how they vary.
Some trees at the investigated site are probably derivatives of a persistent on-site lineage of ancient ancestry, perhaps going back millions of years. It is suggested that this population and additional ones within the southern Dawson Range of Yukon Beringia have potential to advance knowledge about gišogenetic variation and evolution.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f14040787/s1, Figure S1: Mushroom encounters; Table S1: East Fork plants; Table S2: Soil Moisture and pH Values; Ring-width (mm) spreadsheets: DATA1311, DATA1316, DATA1320, DATA1326, DATA1328, DATA1329.
Author Contributions
Conceptualization, R.A.S.; methodology, R.A.S.; validation, R.A.S., X.Y. and H.F.; formal analysis, R.A.S.; investigation, R.A.S., X.Y. and H.F.; resources, R.A.S.; data curation, X.Y., R.A.S. and H.F.; writing—original draft preparation, R.A.S.; writing—review and editing, R.A.S.; visualization, R.A.S. and X.Y.; supervision, R.A.S.; project administration, R.A.S.; funding acquisition, R.A.S. All authors have read and agreed to the published version of the manuscript.
Funding
Funding for 2007–2013 field investigations was from the Government of Canada Program for International Polar Year sponsored by the International Council for Science and the World Meteorological Organization in support of project number 2006-SR1-CC-027, PPS Arctic Canada.
Data Availability Statement
Full data sets are being provided as supplemental files.
Acknowledgments
We are indebted to Petra Foerster for collection and inventory of plants, to Joe Muff for general field assistance, to Grace Wheeler and Bruce Wheeler (deceased) for assistance in shelter. Dean McCarthy, Len Lanteigne, James Farrell and the facilities in the Stem Analysis Lab at the Canadian Forest Service assisted in preparation and scanning of disks in the laboratory. Joanne MacDonald assisted X.Y. in drafting her M.Sc.F. thesis entitled ‘Growth Patterns of Timberline Picea glauca on a site in the Yukon Plateau: Correlations Within Tree and Between Trees,’ University of New Brunswick, 2012.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Savidge, R.A. Tree growth and wood quality. In Wood Quality and Its Biological Basis; Barnett, J.R., Jeronimidis, G., Eds.; CRC Press: Baton Rouge, FL, USA, 2003; Chapter 1; ISBN 0-8493-2819-5. [Google Scholar]
- Gattia, R.C.; Reich, P.B.; Gamarra, J.G.; Crowther, T.; Hui, C.; Morera, A.; Bastin, J.F.; De-Miguel, S.; Nabuurs, G.J.; Svenning, J.C.; et al. The number of tree species on Earth. Proc. Natl. Acad. Sci. USA 2022, 119, e2115329119. [Google Scholar] [CrossRef]
- Lee, Z.H.; Hirakawa, T.; Yamaguchi, N.; Ito, T. The roles of plant hormones and their interactions with regulatory genes in determining meristem activity. Int. J. Mol. Sci. 2019, 20, 4065. [Google Scholar] [CrossRef] [Green Version]
- Savidge, R.A.; Farrar, J.L. Cellular adjustments in the vascular cambium leading to spiral-grain formation in conifers. Can. J. Bot. 1984, 62, 2872–2879. [Google Scholar] [CrossRef]
- Dalessandro, G.; Roberts, L.W. Induction of xylogenesis in pith parenchyma expiants of Lactuca. Am. J. Bot. 1971, 58, 378–385. [Google Scholar] [CrossRef]
- Pinches, T.G. The Babylonian Tablets of the Berens Collection; Asiatic Society Monographs; Royal Asiatic Society: London, UK, 1915; Volume XVI. [Google Scholar]
- Savidge, R.A. Intrinsic regulation of cambial growth. J. Plant Growth Regul. 2001, 20, 52–77. [Google Scholar] [CrossRef]
- Savidge, R.A. Electromagnetic survey report—Mt. Nansen project #158. In Mount Nansen Project #158, Progress Report for Period June 1–30, 1971; Dickinson, R.A., Savidge, R.A., Eds.; Energy, Mines and Resources Property file Collection, ARMC012150; Government of Yukon: Whitehorse, Yukon. Available online: https://data.geology.gov.yk.ca/Reference/72584 (accessed on 30 March 2023).
- Savidge, R.A. Yukon taiga—Past, present and future. In The Biosphere; Ishwaran, N., Ed.; InTech Publisher: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Savidge, R.A. Porsild spruce in Canada—An update. For. Chron. 2013, 90, 77–79. [Google Scholar] [CrossRef] [Green Version]
- Rainville, R.A.; Gajewski, K. Holocene environmental history of the Aishihik Region, Yukon, Canada. Can. J. Earth Sci. 2013, 50, 397–405. [Google Scholar] [CrossRef]
- Anderson, L.L.; Hu, F.H.; Nelson, D.M.; Petit, R.J.; Paige, K.N. Ice-age endurance: DNA evidence of a white spruce refugium in Alaska. Proc. Natl. Acad. Sci. USA 2006, 103, 12447–12450. [Google Scholar] [CrossRef] [Green Version]
- Savidge, R.A. Evidence for early glaciation of southeastern Beringia. Can. J. Earth Sci. 2020, 57, 199–226. [Google Scholar] [CrossRef]
- Smith, C.A.S.; Meikle, J.C.; Roots, C.F. Ecoregions of the Yukon Territory: Biophysical Properties of Yukon Landscapes; Agriculture and Agri-Food Canada, PARC Technical Bulletin No. 04-01; Minister of Supply and Services Canada: Summerland, BC, Canada, 2004; 313p, ISBN 0-660-18828-7. [Google Scholar]
- Jackson, L.E., Jr.; Barendregt, R.W.; Baker, J.; Irving, E. Early Pleistocene volcanism and glaciation in central Yukon: A new chronology from field studies and paleomagnetism. Can. J. Earth Sci. 1996, 33, 904–916. [Google Scholar] [CrossRef]
- LeBarge, W.P. Sedimentology of Placer Gravels Near Mt. Nansen, Central Yukon Territory; Bulletin 4; Exploration and Geological Services Division, Northern Affairs Program; Government of Yukon: Whitehorse, Yukon, 1995; 155p, ISBN 0-662-22923-1.
- Michelsen, A.; Schmidt, I.K.; Jonasson, S.; Dighton, J.; Jones, H.E.; Callaghan, T.V. Inhibition of growth, and effects on nutrient uptake of arctic graminoids by leaf extracts—Allelopathy or resource competition between plants and microbes? Oecologia 1995, 103, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Booysen, M.; Sikes, D.; Bowser, M.; Andrews, R. Earthworms (Oligochaeta: Lumbricidae) of interior Alaska. Biodivers. Data J. 2018, 6, e27427. [Google Scholar] [CrossRef] [PubMed]
- Preece, S.J.; Westgate, J.A.; Froese, D.G.; Pearce, N.J.G.; Perkins, W.T. A catalogue of late Cenozoic tephra beds in the Klondike goldfields and adjacent areas, Yukon Territory. Can. J. Earth Sci. 2011, 48, 1386–1418. [Google Scholar] [CrossRef]
- Shah, R.; Mir, B.A. The freezing point of soils and the factors affecting its depression. In Advances in Construction Management, Lecture Notes in Civil Engineering; Loon, L.Y., Subramaniyan, M., Gunasekaran, K., Eds.; Springer: Singapore, 2022; Volume 191, pp. 157–166. [Google Scholar] [CrossRef]
- Government of Yukon, Wildland Fire Management Branch. Yukon Fire History. Available online: https://mapservices.gov.yk.ca/arcgis/rest/services/GeoYukon/GY_EmergencyManagement/MapServer/14 (accessed on 31 January 2023).
- Savidge, R.A. The tracheid differentiation factor of conifer needles. Int. J. Plant Sci. 1994, 155, 272–290. [Google Scholar] [CrossRef]
- Savidge, R.A. Cell biology of bordered-pit formation in balsam-fir trees. Botany 2014, 92, 495–511. [Google Scholar] [CrossRef]
- Matthews, J.A.; Briffa, K.R. The ‘Little Ice Age’: Re-evaluation of an evolving concept. Geogr. Ann. Ser. A Phys. Geogr. 2005, 87, 17–36. [Google Scholar] [CrossRef]
- Bradley, R.S.; Jones, P.D. ‘Little Ice Age’ summer temperature variations: Their nature and relevance to recent global warming trends. Holocene 1993, 3, 367–376. [Google Scholar] [CrossRef]
- Hamilton, J.A.; El Kayal, W.; Hart, A.T.; Runcie, D.E.; Arango-Velez, A.; Cooke, J.E.K. The joint influence of photoperiod and temperature during growth cessation and development of dormancy in white spruce (Picea glauca). Tree Physiol. 2016, 36, 1432–1438. [Google Scholar] [CrossRef] [Green Version]
- Esper, J.; Frank, D. Divergence pitfalls in tree-ring research. Clim. Change 2009, 94, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Wilmking, M.; Singh, J. Eliminating the “divergence problem” at Alaska’s northern treeline. Clim. Past Discuss. 2008, 4, 741–759. [Google Scholar] [CrossRef] [Green Version]
- Dearborn, K.D.; Danby, R.K. Climatic drivers of tree growth at tree line in southwest Yukon change over time and vary between landscapes. Clim. Change 2018, 150, 211–225. [Google Scholar] [CrossRef]
- Szeicz, J.M.; MacDonald, G.M. Recent white spruce dynamics at the subarctic alpine treeline of northwestern Canada. J. Ecol. 1995, 83, 873–885. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Fastie, C.L. Spatial and temporal variability in the growth and climate response of treeline trees in Alaska. Clim. Change 2002, 52, 481–509. [Google Scholar] [CrossRef]
- Gryc, V.; Hacura, J.; Vavrcík, H.; Urban, J.; Gebauer, R. Monitoring of xylem formation in Picea abies under drought stress influence. Dendrobiology 2012, 67, 15–24. [Google Scholar]
- Young-Robertson, J.M.; Ogle, K.; Welker, J.M. Thawing seasonal ground ice: An important water source for boreal forest plants in Interior Alaska. Ecohydrology 2017, 10, e1796. [Google Scholar] [CrossRef]
- Wang, W.; Hoch, G. Negative effects of low root temperatures on water and carbon relations in temperate tree seedlings assessed by dual isotopic labelling. Tree Physiol. 2022, 42, 1311–1324. [Google Scholar] [CrossRef] [PubMed]
- Savidge, R.A. Climate change, forest mortality, and the need for a solid scientific foundation in forestry. Environ. Sci. Proc. 2022, 22, 44. [Google Scholar] [CrossRef]
- Risk, C.; McKenney, D.W.; Pedlar, J.; Lu, P. A compilation of North American tree provenance trials and relevant historical climate data for seven species. Sci. Data 2021, 8, 29. [Google Scholar] [CrossRef]
- Morgenstern, K.; D’Eon, S.; Penner, M. White spruce growth to age 44 in a provenance test at the Petawawa Research Forest. For. Chron. 2006, 82, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, L.M. A Range-Wide Common Garden Experiment of White Spruce Indicates Population Differentiation in Drought Tolerance Traits. Master’s Thesis, University of Alberta, Edmonton, AB, Canada, 2009. [Google Scholar]
- Prud’homme, G.O.; Lamhamedi, M.S.; Benomar, L.; Rainville, A.; DeBlois, J.; Bousquet, J.; Beaulieu, J. Ecophysiology and growth of white spruce seedlings from various seed sources along a climatic gradient support the need for assisted migration. Front. Plant Sci. 2018, 8, 2214. [Google Scholar] [CrossRef] [Green Version]
- Sebastian-Azcona, J.; Hamann, A.; Hacke, U.G.; Rweyongeza, D. Survival, growth and cold hardiness tradeoffs in white spruce populations: Implications for assisted migration. For. Ecol. Mgmt. 2019, 433, 544–552. [Google Scholar] [CrossRef]
- Szeicz, J.; MacDonald, G. Dendroclimatic reconstruction of summer temperatures in northwestern Canada since A.D. 1638 based on age-dependent modeling. Quat. Res. 1995, 44, 257–266. [Google Scholar] [CrossRef]
- Anderson, L.L.; Hu, F.S.; Paige, K.N. Phylogeographic history of white spruce during the Last Glacial Maximum: Uncovering cryptic refugia. J. Hered. 2011, 102, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lait, L.A.; Burg, T.M. When east meets west: Population structure of a high-latitude resident species, the boreal chickadee (Poecile hudsonicus). Heredity 2013, 111, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Nienstaedt, H.; Zasada, J.C. Picea glauca. In Silvics of North America; Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 204–226. [Google Scholar]
- Alfieri, F.J.; Evert, R.F. Seasonal development of the secondary phloem in Pinus. Am. J. Bot. 1968, 55, 518–528. [Google Scholar] [CrossRef]
- Rainer-Lethaus, G.; Oberhuber, W. Phloem girdling of Norway spruce alters quantity and quality of wood formation in roots particularly under drought. Front. Plant Sci. 2018, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Savidge, R.A. Inductive effects of conifer needles on xylogenesis—Auxin not the explanation. In Cell & Molecular Biology of Wood Formation; Savidge, R.A., Barnett, J.R., Napier, R., Eds.; BIOS Scientific: Oxford, UK, 2000; pp. 237–254. [Google Scholar]
- Hanlon, V.C.T.; Otto, S.P.; Aitken, S.N. Somatic mutations substantially increase the per-generation mutation rate in the conifer Picea sitchensis. Evol. Lett. 2019, 3–4, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Hartig, T. Uber die Entwicklung des Jahresringes des Holzplanzen. Bot. Zeitung 1853, 11, 553–579. [Google Scholar]
- Jost, L. Uber Beziehungen zwischen der Blattentwicklung und del’ Gerassbildun~ in del’ Pflanze. Bot. Zeitung 1893, 51, 89–138. [Google Scholar]
- Went, F. Wuchsstoff und Wachstum. Rec. Trav. Bot. Neerl. 1928, 25, 1–116. [Google Scholar]
- Arteca, R.N. Historical Aspects and Fundamental Terms and Concepts. In Plant Growth Substances; Springer: Boston, MA, USA, 1996. [Google Scholar] [CrossRef]
- Enders, T.A.; Strader, L.C. Auxin activity: Past, present, and future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Napier, R.M. Regulators of Growth | Auxins. In Encyclopedia of Applied Plant Sciences; Thomas, B., Ed.; Elsevier: Amsterdam, The Netherlands, 2003; pp. 985–995. ISBN 9780122270505. [Google Scholar] [CrossRef]
- Savidge, R.A. Regulation of Seasonal Cambial Activity and Tracheid Differentiation in Pinus contorta Dougl. Ph.D. Thesis, University of Wales, Wales, UK, 1981. Available online: www.researchgate.net/publication/318541571 (accessed on 30 March 2023).
- Savidge, R.A.; Wareing, P.F. Plant-growth regulators and the differentiation of vascular elements. In Xylem Cell Development; Barnett, J.R., Ed.; Castle House Publisher: Tunbridge Wells, UK, 1981; pp. 192–235. ISBN 0 7194 0050 3. [Google Scholar]
- Savidge, R.A.; Wareing, P.F. Seasonal cambial activity and xylem development in Pinus contorta in relation to endogenous indol-3-ylacetic acid and (S)-abscisic acid levels. Can. J. For. Res. 1984, 14, 676–682. [Google Scholar] [CrossRef]
- Savidge, R.A.; Wareing, P.F. Apparent auxin production and transport during winter in the non-growing pine tree. Can. J. Bot. 1982, 60, 681–691. [Google Scholar] [CrossRef]
- Oribe, Y.; Kubo, T. Effect of heat on cambial reactivation during winter dormancy in evergreen and deciduous conifers. Tree Physiol. 1997, 17, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Gričar, J.; Zupančič, M.; Čufar, K.; Koch, G.; Schmitt, U.; Oven, P. Effect of local heating and cooling on cambial activity and cell differentiation in the stem of Norway spruce (Picea abies). Ann. Bot. 2006, 97, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Kudo, K.; Nabeshima, E.; Begum, S.; Yamagishi, Y.; Nakaba, S.; Oribe, Y.; Yasue, K.; Funada, R. The effects of localized heating and disbudding on cambial reactivation and formation of earlywood vessels in seedlings of the deciduous ring-porous hardwood, Quercus serrata. Ann. Bot. 2014, 113, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, A.; Mattana, S.; Emiliani, G.; Anichini, M.; Traversi, M.L.; Pavone, F.S.; Cicchi, R. Localized stem heating from the rest to growth phase induces latewood-like cell formation and slower stem radial growth in Norway spruce saplings. Tree Physiol. 2022, 42, 1149–1163. [Google Scholar] [CrossRef]
- Kijidani, Y.; Wu, Z.; Savidge, R. New insight into phytohormone regulation of wood formation in conifers. In Phytomorphology Golden Jubilee Issue 2001: Trends in Plant Science; Rangaswamy, N.S., Ed.; International Society of Plant Morphologists, University of Delhi: Delhi, India, 2001; 586p, pp. 185–200. [Google Scholar]
- Zajaczkowski, S. Auxin stimulation of cambial activity in Pinus silvestris I. The differential cambial response. Physiol. Plant. 1973, 29, 281–287. [Google Scholar] [CrossRef]
- Savidge, R.A. In vitro wood formation in ‘chips’ from merchantable stem regions of Larix laricina. IAWA J. 1993, 14, 3–11. [Google Scholar]
- Fukuda, H.; Komamine, A. Direct evidence for cytodifferentiation to trachearv clements without intervening mitosis in a culture of single cells isolated from the mesophyll of Zinnia elegans. Plant Physiol. 1980, 65, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Savidge, R.A. Auxin and ethylene regulation of diameter growth in trees. Tree Physiol. 1988, 4, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Muday, G.K.; Rahman, A.; Binder, B.M. Auxin and ethylene: Collaborators or competitors? Trends Plant Sci. 2012, 17, 181–195. [Google Scholar] [CrossRef]
- Savidge, R.A. ACC induction of xylogenesis in horseradish roots. Plant Growth Reg. Soc. Am. 2007, 33, 60–65. Available online: www.researchgate.net/publication/311654362 (accessed on 30 March 2023).
- Björklund, S.; Antti, H.; Uddestrand, I.; Moritz, T.; Sundberg, B. Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 2007, 52, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Immanen, J.; Nieminen, K.; Smolander, O.-P.; Kojima, M.; Serra, J.A.; Koskinen, P.; Zhang, J.; Elo, A.; Mähönen, A.P.; Street, N.; et al. Cytokinin and auxin display distinct but interconnected distribution and signaling profiles to stimulate cambial cctivity. Curr. Biol. 2016, 26, 1990–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liua, C.; Yua, H.; Rao, X.; Li, L.; Dixon, R.A. Abscisic acid regulates secondary cell-wall formation and lignin deposition in Arabidopsis thaliana through phosphorylation of NST1. Proc. Natl. Acad. Sci. USA 2021, 118, e2010911118. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.; Park, S.-G.; Hwang, H.; Yoo, S.; Bae, W.; Kim, E.; Kim, J.; Lee, H.-Y.; Heo, T.-Y.; et al. Brassinosteroid-BZR1/2-WAT1 module determines the high level of auxin signalling in vascular cambium during wood formation. New Phytol. 2021, 230, 1503–1516. [Google Scholar] [CrossRef]
- Hu, J.; Hu, X.; Yang, Y.; He, C.; Hu, J.; Wang, X. Strigolactone signaling regulates cambial activity through repression of WOX4 by transcription factor BES1. Plant Physiol. 2022, 188, 255–267. [Google Scholar] [CrossRef]
- Behr, M.; Lutts, S.; Hausman, J.-F.; Guerriero, G. Jasmonic acid to boost secondary growth in hemp hypocotyl. Planta 2018, 248, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Morales-Quintana, L.; Ramos, P. A talk between flavonoids and hormones to reorient the growth of gymnosperms. Int. J. Mol. Sci. 2021, 22, 12630. [Google Scholar] [CrossRef]
- Wong, W.S.; Guo, D.; Wang, X.L.; Yin, Z.Q.; Xia, B.; Li, N. Study of cis-cinnamic acid in Arabidopsis thaliana. Plant Physiol. Biochem. 2005, 43, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Tejos, R.; Beck, M.; Himschoot, E.; Li, H.; Robatzek, S.; Vanneste, S.; Friml, J. Salicylic acid interferes with clathrin-mediated endocytic protein trafficking. Proc. Natl. Acad. Sci. USA 2013, 110, 7946–7951. [Google Scholar] [CrossRef] [Green Version]
- Grana, E.; Costas-Gil, A.; Longueira, S.; Celeiro, M.; Teijeira, M.; Reigosa, M.J.; Sanchez-Moreiras, A.M. Auxin-like effects of the natural coumarin scopoletin on Arabidopsis cell structure and morphology. J. Plant Physiol. 2017, 218, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Savidge, R.A. Dihydroconiferyl alcohol in developing xylem of Pinus contorta. Phytochemistry 1987, 26, 93–94. [Google Scholar] [CrossRef]
- Binns, A.N.; Chen, R.H.; Wood, H.N.; Lynn, D.G. Cell division promoting activity of naturally occurring dehydrodiconiferyl glucosides: Do cell wall components control cell division? Proc. Natl. Acad. Sci. USA 1987, 84, 980–984. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Motose, H.; Iwamoto, K.; Fukuda, H. Expression and genome-wide analysis of the xylogen-type gene family. Plant Cell Physiol. 2011, 52, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Miao, Y.; Huang, H.; Zhang, Y.; Huang, L.; Cao, J. Arabinogalactan proteins: Focus on the role in cellulose synthesis and deposition during plant cell wall biogenesis. Int. J. Mol. Sci. 2022, 23, 6578. [Google Scholar] [CrossRef]
- Fukuda, H.; Hardtke, C.S. Peptide signaling pathways in vascular differentiation. Plant Physiol. 2020, 182, 1636–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soumelidou, K.; Li, H.; Barnett, J.R.; John, P.; Battey, N.H. The effect of auxin and calcium antagonists on tracheary element differentiation in Phaseolus vulgaris L. J. Plant Physiol. 1994, 143, 717–721. [Google Scholar] [CrossRef]
- DeRose, R.J.; Bekker, M.F.; Long, J.N. Traumatic resin ducts as indicators of bark beetle outbreaks. Can. J. For. Res. 2017, 47, 1168–1174. [Google Scholar] [CrossRef]
- Martin, D.; Tholl, D.; Gershenzon, J.; Bohlmann, J. Methyl jasmonate induces traumatic resin ducts, terpenoid resin biosynthesis, and terpenoid accumulation in developing xylem of Norway spruce stems. Plant Physiol. 2002, 129, 1003–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubler, H. Mechanism of frost crack formation in trees—A review and synthesis. For. Sci. 1983, 29, 559–568. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Savidge, R.A.; Yuan, X.; Foerster, H. Gišogenetic Variation in White-Spruce (Picea glauca (Moench) Voss) Trees of Yukon Beringia, Canada. Forests 2023, 14, 787. https://doi.org/10.3390/f14040787
AMA Style
Savidge RA, Yuan X, Foerster H. Gišogenetic Variation in White-Spruce (Picea glauca (Moench) Voss) Trees of Yukon Beringia, Canada. Forests. 2023; 14(4):787. https://doi.org/10.3390/f14040787
Chicago/Turabian StyleSavidge, Rodney Arthur, Xin Yuan, and Hartmut Foerster. 2023. "Gišogenetic Variation in White-Spruce (Picea glauca (Moench) Voss) Trees of Yukon Beringia, Canada" Forests 14, no. 4: 787. https://doi.org/10.3390/f14040787
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.