
Prospects for the Preservation of the Main Pinus sylvestris L. Ecotypes in Poland in the Context of the Habitat Conditions of Their Occurrence


Abstract
:1. Introduction
2. Materials and Methods
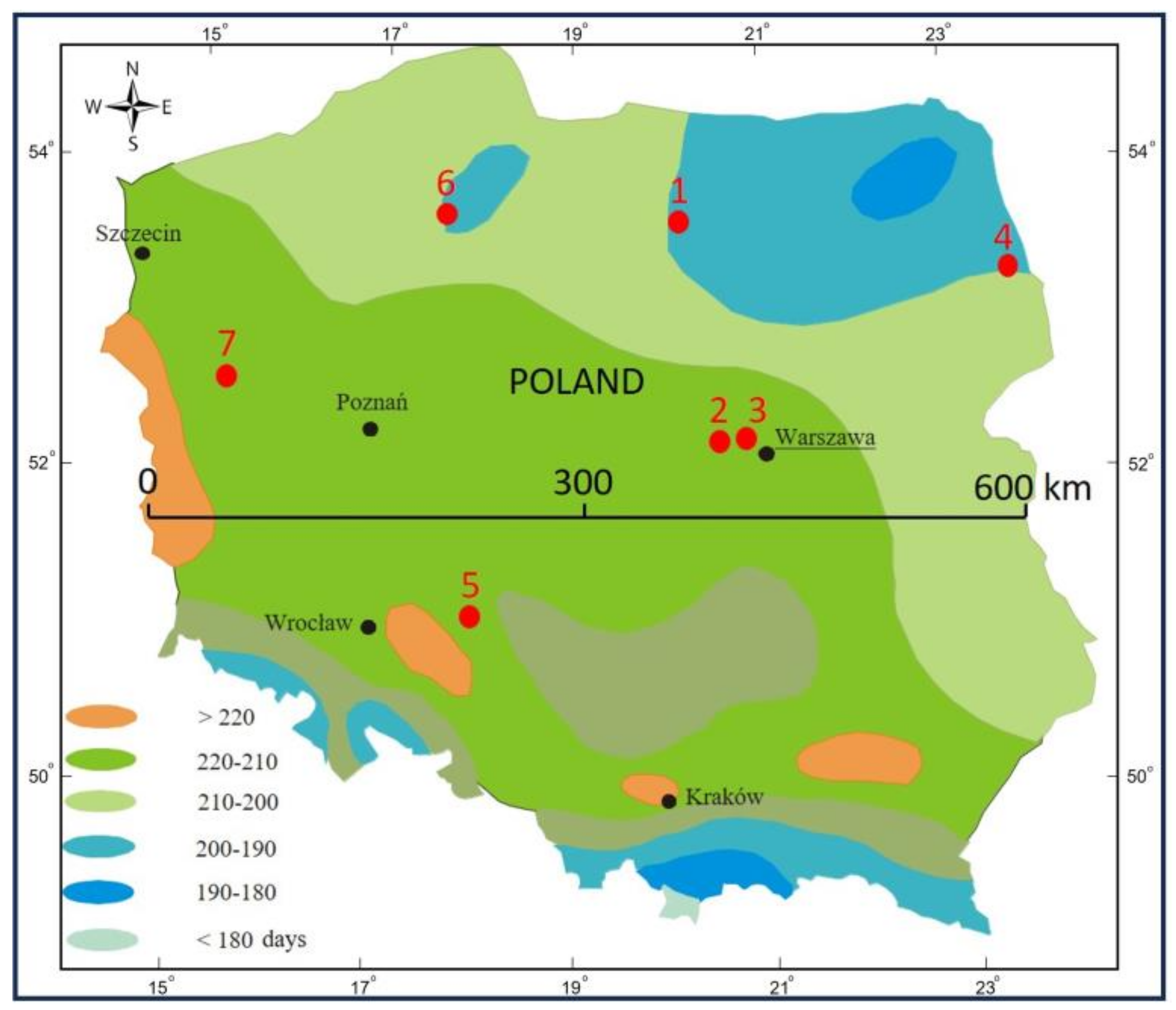
2.1. Research Area
2.2. Calculation of the NDVI
2.3. Statistical Analysis
3. Results
- Tree Slenderness Coefficient (TSC): 1—lowest slenderness coefficient, 7—highest slenderness coefficient; the lower the TSC, the greater the resistance to wind damage.
- Plant community: 1—Leucobryo–Pinetum, 2—Querco roboris–Pinetum, 3—Seratulo–Pinetum, 4—Calamagrostio arundinaceae–Quercetum petraeae, 5—Galio sylvatici–Carpinetum; the lower the index, the higher the likelihood of natural pine regeneration.
- NDVI: 1—lowest median value for the NDVI, 7—highest median value for the NDVI; the higher the NDVI value, the better the habitat conditions and, therefore, the lower the chance of natural pine regeneration due to competition from deciduous tree species.
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Durrant, T.H.; de Rigo, D.; Caudullo, G. Pinus sylvestris in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Przybylski, P.; Konatowska, M.; Jastrzębowski, S.; Tereba, A.; Mohytych, V.; Tyburski, Ł.; Rutkowski, P. The Possibility of Regenerating a Pine Stand through Natural Regeneration. Forests 2021, 12, 1055. [Google Scholar] [CrossRef]
- Lorenc, F.; Véle, A. Characteristics of Pinus sylvestris stands infected by Viscum album subsp. austriacum. Austrian J. For. Sci. 2022, 139, 31–50. Available online: https://www.forestscience.at/content/dam/holz/forest-science/2022/01/CB2201_Art2.pdf (accessed on 12 September 2023).
- Miścicki, S. Changes in the stands of the Białowieża National Park from 2000 to 2015. For. Res. Pap. 2016, 77, 371–379. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Change Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Daszkiewicz, P.; Oleksyn, J. Introduction of the ”Riga pine” in the 18th and 19th century France. Rocz. Dendrol. 2005, 53, 7–40. [Google Scholar]
- Jelonek, T.; Tomczak, A.; Jakubowski, M.; Grzywiński, W.; Pazdrowski, W.; Remlein, A.; Jurkieiwicz, E. An analysis of the quality and value of timber in pine tree stands representing selected ecotypes of this species in Poland. Ann. Wars. Univ. Life Sci. —SGGW For. Wood Technol. 2016, 95, 20–28. [Google Scholar]
- Núñez, M.R.; Calvo, L. Effect of high temperatures on seed germination of Pinus sylvestris and Pinus halepensis. For. Ecol. Manag. 2000, 131, 183–190. [Google Scholar] [CrossRef]
- Turna, I.; Bilgili, E. Effect of heat on seed germination of Pinus sylvestris and Pinus nigra ssp. pallasiana. Int. J. Wildland Fire 2006, 15, 283–286. [Google Scholar] [CrossRef]
- Núñez, M.R.; Bravo, F.; Calvo, L. Predicting the probability of seed germination in Pinus sylvestris L. and four competitor shrub species after fire. Ann. For. Sci. 2003, 60, 75–81. [Google Scholar] [CrossRef]
- Oleksyn, J.; Tjoelker, M.G.; Reich, P.B. Growth and biomass partitioning of populations of European Pinus sylvestris L. under simulated 50° and 60° N daylengths: Evidence for photoperiodic ecotypes. New Phytol. 1992, 120, 561–574. [Google Scholar] [CrossRef]
- Lesiczka, P.; Pawlaczyk, E.M.; Łabiszak, B.; Urbaniak, L. Variability of Scots pine (Pinus sylvestris L.) called Tabórz pine (Forest District Miłomłyn) expressed in analysis of morphology of needle traits and polymorphism of microsatellite DNA. For. Res. Pap. 2017, 78, 136–148. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecology, The Experimental Analysis of Distribution and Abundanc, 4th ed.; PWN: Warsaw, Poland, 1996. [Google Scholar]
- Barzdajn, W. Wyniki 24−letniego doświadczenia proweniencyjnego z sosną Nadleśnictwie Supraśl. Sylwan 2008, 4, 21–29. [Google Scholar]
- Młynarczyk, A.; Konatowska, M.; Królewicz, S.; Rutkowski, P.; Piekarczyk, J.; Kowalewski, W. Spectral Indices as a Tool to Assess the Moisture Status of Forest Habitats. Remote Sens. 2022, 14, 4267. [Google Scholar] [CrossRef]
- Wójkiewicz, B.; Żukowska, W.B.; Urbaniak, L.; Kowalczyk, J.; Litkowiec, M.; Lewandowski, A. Określenie pochodzenia wyłączonych drzewostanów nasiennych sosny rychtalskiej (Pinus sylvestris L.) z wykorzystaniem markerów mikrosatelitarnych. Sylwan 2019, 163, 637–644. [Google Scholar] [CrossRef]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski; PWN: Warsaw, Poland, 2006; p. 537. [Google Scholar]
- Muñoz Sabater, J. ERA5-Land Monthly Averaged Data From 1981 to Present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). 2019. Available online: https://cds.climate.copernicus.eu/cdsapp#!/dataset/reanalysis-era5-land-monthly-means?tab=overview (accessed on 12 September 2023).
- Biały, K.; Brożek, S.; Chojnicki, J.; Czepińska-Kamińska, D.; Januszek, K.; Kowalkowski, A.; Krzyżanowski, A.; Okołowicz, M.; Sienkiewicz, A.; Skiba, S.; et al. Klasyfikacja Gleb Leśnych Polski; CILP: Warsaw, Poland, 2000; p. 125. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022; p. 236. [Google Scholar]
- Kelly, D.L.; Connolly, A. A review of the plant communities associated with Scots pine (Pinus sylvestris L.) in Europe, and an evaluation of putative indicator/specialist species. For. Syst. 2000, 9, 15–39. [Google Scholar] [CrossRef]
- Konatowska, M.; Rutkowski, P. Porównanie Zbiorowisk Roślinnych z Połowy XX i Początku XXI Wieku w Nadleśnictwie Doświadczlnym Zielonka; Publishing House of the University of Life Science: Poznań, Poland, 2021. [Google Scholar]
- Szeligowski, H.; Buraczyk, W.; Konecka, A.; Studnicki, M.; Drozdowski, S. A multi-trait assessment of selected provenances of Scots pine following 50 years of growth on a provenance experiment in Central Poland, in the light of climate change. Eur. J. For. Res. 2023, 142, 509–520. [Google Scholar] [CrossRef]
- Remlein, A.; Jelonek, T.; Tomczak, A.; Jakubowski, M.; Grzywinski, W. Morphological architecture of different ecotypes of Scots pine (Pinus sylvestris L.) in Poland. Ann. Wars. Univ. Life Sci.—SGGW Wood Technol. 2015, 92, 355–363. [Google Scholar]
- Jarmuł, M.; Kaczmarski, Z. Slenderness of Scots pine (Pinus sylvestris L.) according to biosocial position of the tree (Kształtowanie się smukłości sosny pospolitej (Pinus sylvestris L.) w zależności od stanowiska biosocjalnego drzewa). Agron. Sci. 2020, 75, 105–115. [Google Scholar] [CrossRef]
- Ige, P.O.; Komolafe, O.O. Non-linear tree crown ratio models in International Institute of Tropical Agriculture Forest, Ibadan. J. Res. For. Wildl. Environ. 2022, 14, 27–42. Available online: https://www.ajol.info/index.php/jrfwe/article/view/224757 (accessed on 12 September 2023).
- Grulke, N.; Maxfield, J.; Riggan, P.; Schrader-Patton, C. Pre-Emptive Detection of Mature Pine Drought Stress Using Multispectral Aerial Imagery. Remote Sens. 2020, 12, 2338. [Google Scholar] [CrossRef]
- Gallardo-Salazar, J.; Carrillo, A.D.; Pompa-García, M.; Aguirre-Salado, C. Multispectral indices and individual-tree level attributes explain forest productivity in a pine clonal orchard of Northern Mexico. Geocarto Int. 2021, 37, 4441–4453. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Viana-Soto, A.; Camarero, J.J.; Zavala, M.A.; Tijerín, J.; García, M. Using spectral indices as early warning signals of forest dieback: The case of drought-prone Pinus pinaster forests. Sci. Total Environ. 2021, 793, 148578. [Google Scholar] [CrossRef] [PubMed]
- Mantas, V.; Fonseca, L.; Baltazar, E.; Canhoto, J.; Abrantes, I. Detection of Tree Decline (Pinus pinaster Aiton) in European Forests Using Sentinel-2 Data. Remote Sens. 2022, 14, 2028. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No | Name | Coordinates of the Middle of Research Plot | Age (2022) | Area (ha) |
|---|---|---|---|---|
| 1 | Taborska Pine | 53°46′30.8″ N; 20°01′32.1″ E | 268 | 15.20 |
| 2 | Kampinoski NP-01 | 52°20′26.3″ N; 20°23′41.8″ E | 188 | 19.05 |
| 3 | Kampinoski NP-02 | 52°20′12.3″ N; 20°46′33.3″ E | 183 | 12.59 |
| 4 | Supraska Pine | 53°15′08.3″ N; 23°29′44.9″ E | 196 | 3.14 |
| 5 | Rychtalska Pine | 51°11′18.4″ N; 17°56′55.3″ E | 142 | 15.15 |
| 6 | Bory Tucholskie NP | 53°47′44.3″ N; 17°34′46.1″ E | 140 | 19.28 |
| 7 | Notecka Pine | 52°42′08.0″ N; 15°28′15.1″ E | 133 | 3.03 |
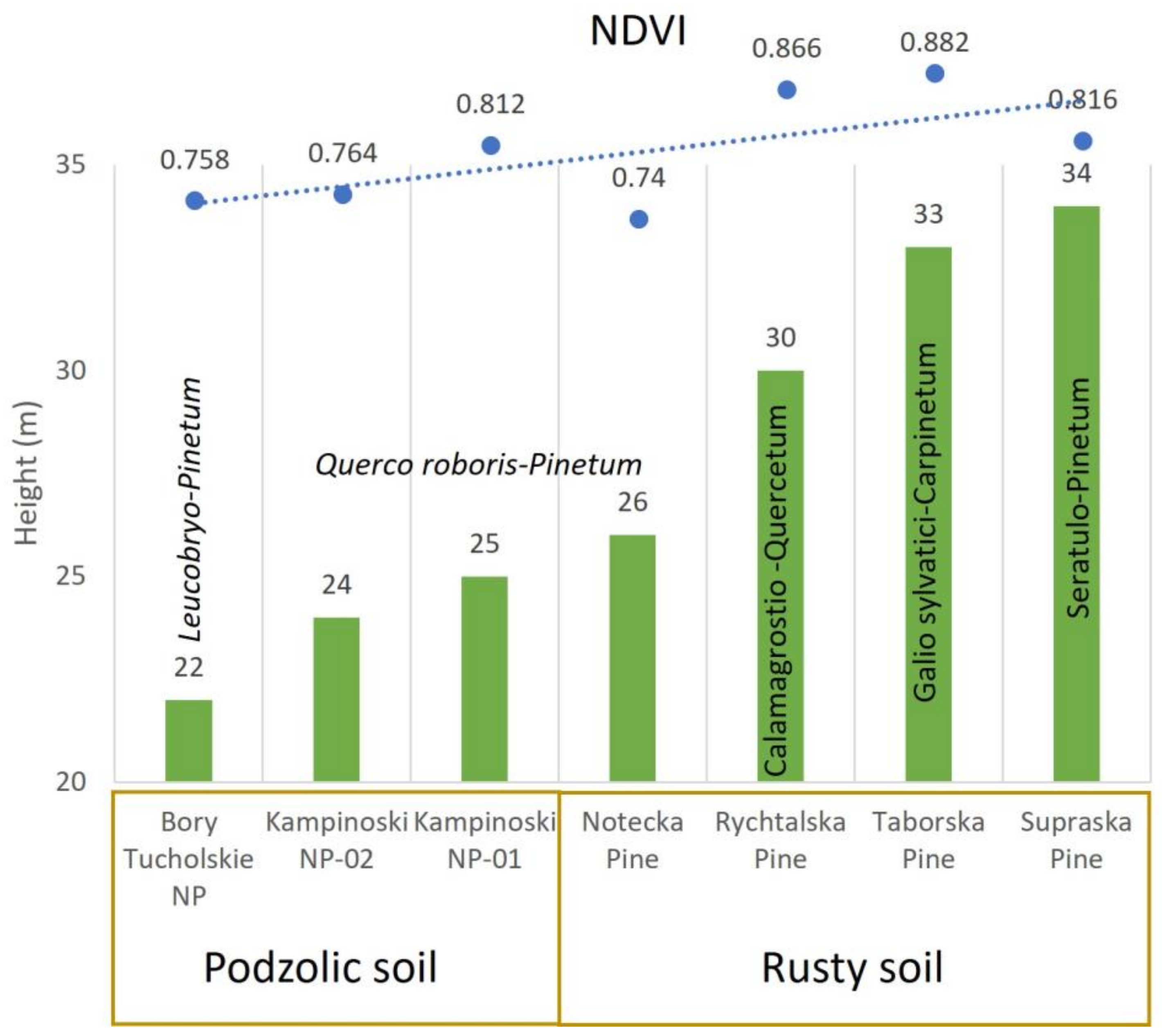
| No | Name | Soil Type | Potential Plant Community | Av. H (m) | Av. DBH (cm) | (H/D) ×100 |
|---|---|---|---|---|---|---|
| 1 | Taborska Pine | PRS * | Eutrophic oak–hornbeam forest (Galio sylvatici–Carpinetum plant community) | 33 | 66 | 50 |
| 2 | Kampinoski NP-01 | PS | Oligotrophic Quercus robur–Pinus sylvestris forest (Querco roboris–Pinetum) | 25 | 45 | 56 |
| 3 | Kampinoski NP-02 | PS | Oligotrophic Quercus robur–Pinus sylvestris forest (Querco roboris–Pinetum) | 24 | 46 | 52 |
| 4 | Supraska Pine | TRS | Mesotrophic Quercus robur–Pinus sylvestris forest (Seratulo–Pinetum) | 34 | 49 | 69 |
| 5 | Rychtalska Pine | BRS | Mesotrophic Quercus petraea forest (Calamagrostio Arundinaceae–Quercetum petraeae) | 30 | 41 | 73 |
| 6 | Bory Tucholskie NP | PS | Oligotrophic pine forest (Leucobryo–Pinetum) | 22 | 33 | 67 |
| 7 | Notecka Pine | PRS | Oligotrophic Quercus robur–Pinus sylvestris forest (Querco roboris–Pinetum) | 26 | 37 | 70 |
| No | Name | Mean | Median | Standard Deviation | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jul. | Aug. | Spt. | Av. | Jul. | Aug. | Spt. | Av. | Jul. | Aug. | Spt. | ||
| 1 | Taborska Pine | 0.794 | 0.793 | 0.793 | 0.793 | 0.885 | 0.881 | 0.880 | 0.882 | 0.262 | 0.261 | 0.261 |
| 2 | Kampinoski NP-01 | 0.822 | 0.832 | 0.784 | 0.812 | 0.823 | 0.828 | 0.785 | 0.812 | 0.013 | 0.021 | 0.012 |
| 3 | Kampinoski NP-02 | 0.760 | 0.787 | 0.758 | 0.768 | 0.756 | 0.785 | 0.750 | 0.764 | 0.016 | 0.014 | 0.025 |
| 4 | Supraska Pine | 0.580 | 0.588 | 0.593 | 0.587 | 0.812 | 0.822 | 0.815 | 0.816 | 0.367 | 0.372 | 0.375 |
| 5 | Rychtalska Pine | 0.885 | 0.853 | 0.859 | 0.866 | 0.888 | 0.851 | 0.859 | 0.866 | 0.019 | 0.014 | 0.013 |
| 6 | Bory Tucholskie NP | 0.788 | 0.808 | 0.682 | 0.759 | 0.789 | 0.808 | 0.677 | 0.758 | 0.012 | 0.010 | 0.019 |
| 7 | Notecka Pine | 0.718 | 0.737 | 0.746 | 0.734 | 0.719 | 0.742 | 0.757 | 0.740 | 0.033 | 0.039 | 0.035 |
| Ecotype Name | Jun. | Jul. | Aug. | Spt. | Av. | Sum | NDVI Median |
|---|---|---|---|---|---|---|---|
| Taborska Pine | 93 | 137 | 29 | 32 | 72.8 | 291 | 0.882 |
| Kampinoski NP-01 | 141 | 62 | 128 | 47 | 94.5 | 378 | 0.812 |
| Kampinoski NP-02 | 99 | 77 | 136 | 51 | 90.8 | 363 | 0.764 |
| Supraska Pine | 71 | 70 | 130 | 24 | 73.8 | 295 | 0.816 |
| Rychtalska Pine | 83 | 58 | 30 | 63 | 58.5 | 234 | 0.866 |
| Bory Tucholskie NP | 50 | 111 | 25 | 28 | 53.5 | 214 | 0.758 |
| Notecka Pine | 26 | 120 | 41 | 37 | 56 | 224 | 0.740 |
| Rank | ||||
|---|---|---|---|---|
| TSC | Plant Community (Competition) | NDVI | Sum | |
| Taborska Pine | 1 | 5 | 7 | 13 |
| Kampinoski NP-01 | 3 | 2 | 4 | 9 |
| Kampinoski NP-02 | 2 | 2 | 3 | 7 |
| Supraska Pine | 5 | 3 | 5 | 13 |
| Rychtalska Pine | 7 | 4 | 6 | 17 |
| Bory Tucholskie NP | 4 | 1 | 2 | 7 |
| Notecka Pine | 6 | 2 | 1 | 9 |
| Parameter | Correlation Coefficients |
|---|---|
| TSC/Plant community | –0.11 |
| TSC/NDVI | –0.21 |
| H/NDVI | 0.74 |
| D/NDVI | 0.70 |
| Plant community/NDVI | 0.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konatowska, M.; Młynarczyk, A.; Rutkowski, P. Prospects for the Preservation of the Main Pinus sylvestris L. Ecotypes in Poland in the Context of the Habitat Conditions of Their Occurrence. Forests 2023, 14, 1967. https://doi.org/10.3390/f14101967
Konatowska M, Młynarczyk A, Rutkowski P. Prospects for the Preservation of the Main Pinus sylvestris L. Ecotypes in Poland in the Context of the Habitat Conditions of Their Occurrence. Forests. 2023; 14(10):1967. https://doi.org/10.3390/f14101967
Chicago/Turabian StyleKonatowska, Monika, Adam Młynarczyk, and Paweł Rutkowski. 2023. "Prospects for the Preservation of the Main Pinus sylvestris L. Ecotypes in Poland in the Context of the Habitat Conditions of Their Occurrence" Forests 14, no. 10: 1967. https://doi.org/10.3390/f14101967