Terpene Production Varies in Pinus thunbergii Parl. with Different Levels of Resistance, with Potential Effects on Pinewood Nematode Behavior


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and PWN Sources
2.2. Resistant and Susceptible P. thunbergii Branches Inoculated with PWN
2.3. Migration, Population Size and Morphological Observations of the PWNs
2.4. Determination of Cellulase Activity Secreted by PWNs
2.5. Real-Time qRT-PCR of Terpene Synthesis-Related Genes
2.6. Terpene Species and Content Determination
2.7. Analysis of Data
3. Results
3.1. Phenotypes of Resistant and Susceptible P. thunbergii after Inoculation with PWN
3.2. Differences in Morphology, Migration and Population Size of PWNs in P. thunbergii with Different Levels of Resistance
3.3. Differences in Cellulase Secretion by PWNs in P. thunbergii with Different Levels of Resistance
3.4. Expression of Terpene Synthesis-Related Genes in P. thunbergii Inoculated with PWNs
3.5. Changes in Terpenoids in P. thunbergii after Inoculation with PWNs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Branco, M.; Bragança, H.; Sousa, E.; Phillips, A.J. Pests and Diseases in Portuguese Forestry: Current and New Threats. In Forest Context and Policies in Portugal; Reboredo, F., Ed.; Springer: Lisbon, Portugal, 2014; Volume 19. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 96, 770–780. [Google Scholar] [CrossRef]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar]
- Togashi, K.; Shigesada, N. Spread of the pinewood nematode vectored by the Japanese pine sawyer: Modeling and analytical approaches. Popul. Ecol. 2006, 48, 271–283. [Google Scholar] [CrossRef]
- National Forestry and Grassland Administration. Announcement of the National Forestry and Grassland Administration (No.6, 2022) (Pine Wood Nematode Disease Epidemic Area in 2022) [N]; Announcement of the National Forestry and Grassland Administration: Beijing, China, 2022. [Google Scholar]
- Choi, W.I.; Nam, Y.; Lee, C.Y.; Choi, B.K.; Shin, Y.J.; Lim, J.-H.; Koh, S.-H.; Park, Y.-S. Changes in Major Insect Pests of Pine Forests in Korea Over the Last 50 Years. Forests 2019, 10, 692. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente, B.; Beck, P.S.A. Management measures to control pine wood nematode spreadin Europe. J. Appl. Ecol. 2019, 56, 2577–2580. [Google Scholar] [CrossRef]
- Liu, Y.; Ponpandian, L.N.; Kim, H.; Jeon, J.; Hwang, B.S.; Lee, S.K.; Park, S.-C.; Bae, H. Distribution and diversity of bacterial endophytes from four Pinus species and their efficacy as biocontrol agents for devastating pine wood nematodes. Sci. Rep. 2019, 9, 12461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wu, X.-Q.; Wang, J. Determination and histopathological observation of resistance to PWD in Pinus densiflora. Natural Sciences Edition. J. Nanjing For. Univ. 2007, 31, 110–114. [Google Scholar]
- Lorio, P.L.; Stephen, F.M.; Paine, T.D. Environment and ontogeny modify loblolly pine response to induced acute water deficits and bark beetle attack. For. Ecol. Manag. 1995, 73, 97–110. [Google Scholar] [CrossRef]
- Lu, F.; Guo, K.; Chen, A.; Chen, S.; Lin, H.; Zhou, X. Transcriptomic profiling of effects of emamectin benzoate on the pine wood nematode Bursaphelenchus xylophilus. Pest Manag. Sci. 2020, 76, 747–757. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Q.; Zhou, Z.; Yin, H.; Wei, Y. Two terpene synthases in resistant Pinus massoniana contribute to defense against Bursaphelenchus xylophilus. Plant Cell Environ. 2020, 44, 257–274. [Google Scholar] [CrossRef]
- Lee, J.-P.; Sekhon, S.S.; Kim, J.H.; Kim, S.C.; Cho, B.-K.; Ahn, J.-Y.; Kim, Y.-H. The Pine Wood Nematode Bursaphelenchus xylophilus and Molecular Diagnostic Methods. Mol. Cell. Toxicol. 2020, 17, 1–13. [Google Scholar] [CrossRef]
- John, T.H.; Radhika, d.; Andrew, C.; Roger, D.H.; Steven, J.N. Cell signalling following plant/pathogen interactions involves the generation of reactive oxygen and reactive nitrogen species. Plant Physiol. Biochem. 2002, 40, 611–617. [Google Scholar]
- Wang, J.; Higgins, V.J. Nitric oxide modulates H2O2-mediated defenses in the Colletotrichum coccodes–tomato interaction. Physiol. Mol. Plant Pathol. 2005, 67, 131–137. [Google Scholar] [CrossRef]
- Keeling, C.I.; Bohlmann, J. Genes, enzymes and chemicals of terpenoid diversity in the constitutive and induced defence of conifers against insects and pathogens. New Phytol. 2006, 170, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Xu, S.; Xu, C.; Lu, H.; Zhang, Z.; Zhang, D.; Mu, W.; Liu, F. Effects of trans-2-hexenal on reproduction, growth and behaviour and efficacy against the pinewood nematode, Bursaphelenchus xylophilus. Pest Manag. Sci. 2016, 73, 888–895. [Google Scholar] [CrossRef]
- Harman-Ware, A.E.; Davis, M.F.; Peter, G.F.; Wang, Y.; Sykes, R.W. Estimation of terpene content in loblolly pine biomass using a hybrid fast-GC and pyrolysis-molecular beam mass spectrometry method. J. Anal. Appl. Pyrolysis 2017, 124, 343–348. [Google Scholar] [CrossRef]
- Manninen, A.M.; Holopainen, M. Variation in Growth, Chemical Defense, and Herbivore Resistance in Scots Pine Provenances. J. Chem. Ecol. 1998, 24, 1315–1331. [Google Scholar] [CrossRef]
- Li, X.-F.; Jiao, H.; Yuan, Y.; Tian, S.-N. Allelopathy and chemical composition analysis of volatile compounds from branches and leaves of Cedrus. Ecol. Environ. Sci. 2015, 24, 263–269. [Google Scholar]
- Dong, G.-P.; Yang, L. Preliminary report on the correlation between volatile odor components of Pinus massoniana and its resistance to PWD. Anhui For. Sci. Technol. 2015, 41, 12–14. [Google Scholar]
- Takeuchi, Y.; Futai, K. Volatile compounds in pine stands suffering from pine wilt disease: Qualitative and quantitative evaluation. Nematology 2006, 8, 869–879. [Google Scholar]
- Ren, Q.; Hu, Y.-J.; Li, Z.-Y.; Jin, Y.-J. Lignin content and peroxidase activity in victimized Pinus massoniana. Acta Ecol. Sin. 2007, 27, 4895–4899. [Google Scholar]
- Sasaki, S.; Odani, K.; Nishiyama, Y. Development and Recovery of Pine Wilt Disease Studied by Tracing Ascending Sap Flow Marked with Water Soluble Stains. J. Jpn. For. Soc. 1987, 66, 141–148. [Google Scholar]
- Wu, X.-Q.; Zhang, Y.; Chen, W.-S. Resistance and histopathological observation of 13 resistant families of Pinus thunbergii to Bursaphelenchus xylophilus. Acta Phytopathol. Sin. 2008, 38, 44–50. [Google Scholar]
- Viglierchio, D.R.; Schmitt, R.V. On the methodology of nematode extraction from field samples: Comparison of methods for soil extraction. J. Nematol. 1983, 15, 450–454. [Google Scholar]
- Yang, X.; Wu, X.Q.; Zhou, A.D.; John, J. Bacterial Diversity and Community Structure in the Pine Wood Nematode Bursaphelenchus xylophilus and B. mucronatus with Different Virulence by High-Throughput Sequencing of the 16S rDNA. PLoS ONE 2015, 10, e0137386. [Google Scholar]
- Kusunoki, M. Symptom Development of Pine Wilt Disease-Histopathological Observations with Electron Microscopes. Jpn. J. Phytopathol. 1987, 53, 622–629. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. In Basic Protein and Peptide Protocols, 1st ed.; Humana Press: Totowa, NJ, USA, 1994; pp. 11–15. [Google Scholar]
- Hu, K.-J.; Yang, B.-J. Studies on enzyme electrophoresis of different strains of Bursaphelenchus xylophilus and Bmucronatus. For. Res. 1995, 1, 73–77. [Google Scholar]
- Jiang, L.-Y.; Wang, X.-Y. Qualitative determination of cellulase in extracts and secretions of Bursaphelenchus xylophilus. For. Pest Dis. 1995, 3, 9–11. [Google Scholar]
- Manninen, A.M.; Tarhanen, S.; Vuorinen, M.; Kainulainen, P. Comparing the Variation of Needle and Wood Terpenoids in Scots Pine Provenances. J. Chem. Ecol. 2002, 28, 211–228. [Google Scholar] [CrossRef]
- Wu, Y.; Cheng, L. Advances in the pathogenesis of PWD. South China Agric. 2018, 6, 73–75. [Google Scholar]
- Li, Y.-X.; Zhang, X.-Y. Progress in pathogenesis of PWD. J. Environ. Entomol. 2018, 40, 231–241. [Google Scholar]
- Kusumoto, D.; Yonemichi, T.; Inoue, H.; Hirao, T.; Watanabe, A.; Yamada, T. Comparison of histological responses and tissue damage expansion between resistant and susceptible Pinus thunbergii infected with pine wood nematode Bursaphelenchus xylophilus. J. For. Res. 2017, 19, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, K. Inhibiting factors of symptom development in several Japanese red pine (Pinus densiflora) families selected as resistant to pine wilt. J. For. Res. 2004, 9, 217–224. [Google Scholar] [CrossRef]
- Xue, M.-J.; Li, Y.; Zhang, J. Relationship between cellulase activity and pathogenicity of Bursaphelenchus mucronatus. J. Biosaf. 2019, 28, 181–188. [Google Scholar]
- Futuyma, D.J.; Rosenthal, G.A.; Janzen, D.H. Herbivores: Their Interactions with Secondary Plant Metabolites. BioScience 1991, 30, 165–219. [Google Scholar]
- Liu, Q.; Wei, Y.; Xu, L.; Hao, Y.; Chen, X.; Zhou, Z. Transcriptomic Profiling Reveals Differentially Expressed Genes Associated with Pine Wood Nematode Resistance in Masson Pine (Pinus massoniana Lamb). Sci. Rep. 2017, 7, 4693. [Google Scholar] [CrossRef]
- Koyama, T.; Ogura, K. Isopentenyl diphosphate isomerase and prenyltransferases. In Isoprenoids, Including Carotenoids and Steroids; Comprehensive natural products chemistry, Elsevier: London, UK, 1999; Volume 2, pp. 69–96. [Google Scholar]
- Zhao, Z.-D.; Li, D.-M.; Hu, X.; Xu, F.-Y. Studies on the Chemical Constituents and resistance Mechanism of Pinus massoniana provenances to pine wood nematode Disease (Report iii). In Studies on the Relationship between the Content of Neutral Terpenes in Pinus massoniana Provenances Resistant to Pine Wood Nematode Inoculation; Chemistry and Industry of Forest Products: Nanjing, China, 2001. [Google Scholar]
- Wu, T.; Liu, J.; Li, M.; Zhang, G.; Liu, L.; Li, X.; Men, X.; Xian, M.; Zhang, H. Improvement of sabinene tolerance of Escherichia coli using adaptive laboratory evolution and omics technologies. Biotechnol. Biofuels 2020, 13, 79–93. [Google Scholar] [CrossRef]
- Cao, B.; Xu, W.-R.; Lu, W. Chemical composition analysis of several insect resistant tree species and study on insect resistance mechanism. For. Sci. Technol. 1998, 6, 27–29. [Google Scholar]
- Bing, C.; Zhong, L.Z.; Long, J.X.; Zeng, X.X. A Study on Scattering Effects of the Extracts from Ailanthus altissima on Anoplophora glabripennis. J. Nanjing For. Univ. 2004, 28, 47–49. [Google Scholar]






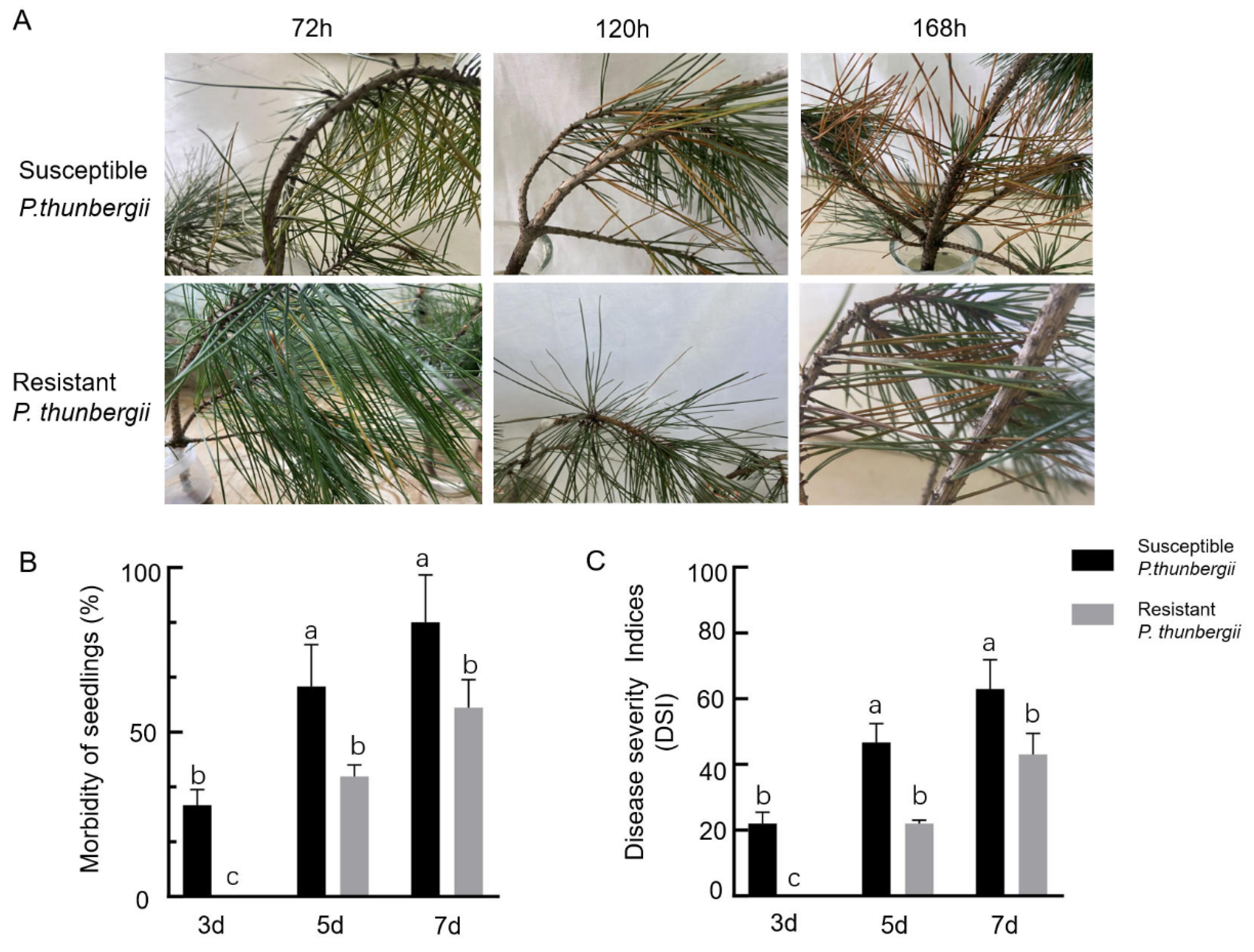{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. thunbergii with Different Levels of Resistance | Distance to Inoculation Site (cm) | |||
|---|---|---|---|---|
| 10 cm above the Inoculation Point | 5 cm above the Inoculation Point | 5 cm below the Inoculation Point | 10 cm below the Inoculation Point | |
| Susceptible P. thunbergii | 670 ± 182 b | 456 ± 141 a | 113 ± 107 a | 50 ± 50 a |
| Resistant P. thunbergii | 160 ± 53 a | 466 ± 161 a | 186 ± 122 a | 25 ± 25 a |
| Table | Relative Percentage Content (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 h | 24 h | 72 h | 168 h | ||||||
| Resistant P. thunbergii | Susceptible P. thunbergii | Resistant P. thunbergii | Susceptible P. thunbergii | Resistant P. thunbergii | Susceptible P. thunbergii | Resistant P. thunbergii | Susceptible P. thunbergii | ||
| 1 | Caryophyllene | 15.71 | 18.4 | 15.06 | 16.8 | 13.12 | 18.87 | 12.82 | 17.36 |
| 2 | γ-Cadinene | 11.46 | 2.89 | 9.82 | - | 5.47 | 2.83 | 3.64 | 2.77 |
| 3 | α-Thujene | 10.82 | - | - | - | 14.14 | - | 9.1 | - |
| 4 | Pseudolimonen | 10.37 | - | 9.27 | - | 7.63 | - | - | - |
| 5 | α-Pinene | 9.61 | 7.93 | 11.77 | - | 15.06 | 15.01 | 16.12 | 16.44 |
| 6 | β-Pinene | 9.61 | 15.27 | 10.43 | - | 14.74 | 21.49 | 16.37 | 19.79 |
| 7 | Tricyclene | 7.85 | 0.11 | 7.15 | 18.78 | 0.12 | 0.3 | 0.1 | 0.18 |
| 8 | Gemacrene D | 6.95 | 8.95 | 6.23 | 8.82 | 5.01 | 9.99 | 7.91 | 10.55 |
| 9 | Terpinlene | 6.94 | 4.13 | 5.95 | 4 | 6.313333 | 4.54 | 6.49 | 3.1 |
| 10 | β-Myrcene | 4.27 | 1.84 | 4.33 | 2.21 | 5.94 | 2.7 | 5.85 | 3.43 |
| 11 | Sabinen | - | - | - | - | 15.97 | - | 18.89 | |
| 12 | β-Cis-ocimene | - | - | - | - | 11.07 | 17.28 | 11.74 | 13.05 |
| P. thunbergii with Different Levels of Resistance | Terpenoids | Inoculation Time | ||
|---|---|---|---|---|
| 0 h | 24 h | 72 h | ||
| Resistant P. thunbergii | cis-γ-Bisabolene | 1.16 | ||
| Linalool | 0.5 | 1.14 | ||
| Alloaromadendrene | 1.73 | 1.46 | ||
| Germacrene B; | 5.38 | 2.71 | ||
| Bicylogermacrene | 4.06 | 0.62 | 0.22 | |
| Dehydrosabinene | 0.63 | 0.59 | 0.43 | |
| Pseudolimonen | 10.37 | 9.27 | 7.63 | |
| α-Thujene | 10.82 | 14.14 | ||
| α-Terpineol | 0.18 | 0.24 | ||
| epi-β-Caryophyllen | 1.21 | |||
| Myrtenal | 0.12 | |||
| Benzene, n-butyl- | 0.37 | |||
| Caryophyllene oxide | 0.12 | |||
| Sabinene | 15.97 | |||
| Susceptible P. thunbergii | (-)-β-Cadinene | 4.39 | 4.7 | |
| α-Muurolene | 2.5 | 0.27 | 0.58 | |
| Borneol | 0.14 | 0.12 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-Y.; Wu, X.-Q.; Wen, T.-Y.; Feng, Y.-Q.; Zhang, Y. Terpene Production Varies in Pinus thunbergii Parl. with Different Levels of Resistance, with Potential Effects on Pinewood Nematode Behavior. Forests 2022, 13, 1140. https://doi.org/10.3390/f13071140
Wang X-Y, Wu X-Q, Wen T-Y, Feng Y-Q, Zhang Y. Terpene Production Varies in Pinus thunbergii Parl. with Different Levels of Resistance, with Potential Effects on Pinewood Nematode Behavior. Forests. 2022; 13(7):1140. https://doi.org/10.3390/f13071140
Chicago/Turabian StyleWang, Xin-Yu, Xiao-Qin Wu, Tong-Yue Wen, Ya-Qi Feng, and Yan Zhang. 2022. "Terpene Production Varies in Pinus thunbergii Parl. with Different Levels of Resistance, with Potential Effects on Pinewood Nematode Behavior" Forests 13, no. 7: 1140. https://doi.org/10.3390/f13071140