
Man-Made Changes in the Flora and Vegetation of Poland: Current Review

Department of Systematic and Environmental Botany, Faculty of Biology, Adam Mickiewicz University in Poznań, Uniwersytetu Poznańskiego 6, 61-614 Poznań, Poland
Diversity 2023, 15(5), 618; https://doi.org/10.3390/d15050618
Submission received: 12 March 2023
/
Revised: 25 April 2023
/
Accepted: 28 April 2023
/
Published: 2 May 2023
(This article belongs to the Special Issue Changes and Evolution of Flora and Vegetation under Human Impacts)
Abstract
:This paper presents the main directions and range of man-made changes to the vascular flora and their effects at the vegetation level. The native flora of Poland, located in the temperate zone of Central Europe, comprises approximately 2639 species. The effect of human pressure, which began in the Neolithic and has intensified in the last 200 years, has caused changes in the native flora and has included an inflow of 975 alien species, 560 of which have become permanently established. Currently, at least 704 native species are recognized as endangered, and only 623 are recognized as being able to occupy habitats that were created or strongly transformed by humans. Native species (apophytes) and aliens (anthropophytes) have co-created completely new plant communities in these habitats. Among the alien plants, neophytes (399 species) outnumber archaeophytes (161 species). Additionally, many neophytes (123 species) can penetrate seminatural and natural communities. Research has shown that the proportions of apophytes, alien plants, and declining species can be used to clearly differentiate plant communities and be used as an indicator of anthropogenic changes. A comparison of the man-made changes in Polish flora and vegetation in other countries showed that the changes in Poland are part of the global process of environmental degradation and are aggravated by the country’s economic development.
1. Introduction
The influence of humans on the natural environment and its components increases with the progress of civilization [1]. The human footprint on the global environment has now become so consequential that it rivals some of the great forces of nature in the magnitude of its influence on the functioning of the Earth system [2]. Today’s cultural landscape is the result of human impact on natural ecosystems over millennia and, in more recent times, the purposeful creation of landscapes, specifically for agricultural production. In densely inhabited regions, i.e., in large parts of Europe, human activity completely masks natural factors. This has applied to Central and Northern Europe since the Middle Ages and to the Mediterranean area since antiquity. Thus, it is very difficult to recognize natural alterations during later periods using the currently available methods of vegetation history [3]. Nevertheless, man-made changes in flora and vegetation have been the subject of regular geobotanical studies for more than 170 years [4,5], and these studies have already yielded many interesting results and hypotheses. The works of Thellung [6,7,8,9] and Linkola [10] were particularly influential in their interpretation as well as the language used to describe the anthropogenic changes of the flora in Europe. The first formed the fundamentals for classifying plants that spread with human influence beyond the limits of their natural range. The second drew attention to the two opposing directional reactions of native species (retreat and spread) and the lack of response to human impact.
In the second half of the 20th century, anthropogenic changes in flora and vegetation became the central focus of geobotanical research, especially in Central Europe, where the effects of human activity were increasingly visible and dramatic. The survey covered a wide range of habitats and plant communities, including forests [11,12], river valleys [13,14], water bodies [15,16], meadows and grasslands [17,18], agricultural fields [19,20], towns [21,22,23,24], rivers [25] and seaports [26], railroads [27,28,29,30], and roadways [31].
Some attempts at obtaining a holistic view of the anthropogenic changes in the flora and the flow and effects of this process were made by Kornaś [32,33,34], Sukopp [35,36], and Faliński [37]. According to Sukopp [35,36], anthropogenic changes in plant cover are manifested at many levels of organization (e.g., populations, species, flora, phytocoenoses) and include two phenomena that are opposed in a dynamic sense: the decline in native species and plant communities and the spread of alien species and the phytocoenoses dominated by them. An important development is the explicit emphasis on the importance of historical factors in the study of anthropogenic changes in flora and vegetation. In 1987, Sukopp and Trepl [38] wrote: “Generalizing and actualistic studies are of value only in an essentially historical theoretical framework. This becomes particularly obvious if one considers two historical processes that have truly revolutionized the structure and function of the actual biotic communities, i.e., the breakdown of barriers between the isolated plant and animal territories that have existed since the Tertiary Period, beginning with the development of worldwide transportation routes around the year 1500 and the rapidly accelerated extermination of species that began in the middle of the 20th century”.
A similar approach to studying the anthropogenic changes of flora and vegetation was represented by Kornaś [32,33,34] and Faliński [37], who proposed the term synanthropization (from Greek syn—together, anthropos—man) to describe this process. Synanthropization, according to Kornaś, is the totality of the contemporary and historical changes in vegetation cover, marked by human activity and primarily affecting the flora and vegetation. Considering the increasing number of research results on the human impact on flora and vegetation, Faliński [37,39] defined synanthropization as part of the directional changes that are found globally as a result of human activity, while pointing out that the key mechanism of this process is the replacement of (i) specific, i.e., endemic, components by nonspecific, i.e., cosmopolitan, components; (ii) native, i.e., autochthonous, components by alien, i.e., allochthonous, elements, and (iii) stenotopic components by eurytopic ones.
These hypotheses have been repeatedly confirmed in several local and regional studies, but due to their poor dissemination, they have rarely been referenced in the discussion. At the turn of the 20th century, research on anthropogenic changes in flora and vegetation was developing rapidly. However, it was going in two independent directions: the first was concerned with the extinction of species, while the second was concerned with the invasion of alien plants. This is confirmed by the dynamics of the papers that were published in journals indexed in the Web of Science database between 1980 and 2022 (Figure 1). Significantly fewer papers have reported a comprehensive analysis of both at the global [40] or regional flora level [41,42].
This paper aimed to present the trends and scope of anthropogenic changes in the vascular flora of Poland, a country highly representative of Central Europe. Using the current knowledge of flora diversity, data on the progress of species extinction, adaptation of native plants to anthropogenic habitats and naturalization of alien species in plant communities in Poland are presented.
1.1. Study Area: The Location and Factors Influencing Both the Flora and the Vegetation
The flora and vegetation of any area are influenced by natural factors related to its geographic location (geology, relief, soils, water conditions, fauna) as well as current and historical anthropogenic factors. A brief description of the study area provides only an outline of the abiotic natural and anthropogenic factors that impact the structure and dynamics of the flora and vegetation of Poland.
Poland is located in Central Europe between the Baltic Sea to the north and the Sudeten and Carpathian Mountains to the south, mostly in the Vistula and Oder River basins. The country’s area is 322,719 square kilometers, making it the 9th largest country in Europe. The inland area covers 311,895 square kilometers, the internal marine waters cover 2041 square kilometers, and the territorial sea covers 8783 square kilometers. Poland’s landscape is dominated by agricultural land, occupying 60% of the area. Approximately 30% of the country is covered by forested and wooded land. Built-up and urbanized land occupies less than 6%, land under water about 2.1%. The remaining area is wasteland [44].
1.1.1. Natural Factors
The study area is dominated by lowland areas (less than 200 meters above the mean sea level), which occupy as much as 75% of the whole area [44,45]. Lowlands are found in the northern and central parts of the country, while mountainous and highland areas are found in the south (Figure 2). Poland is one of several countries in Europe with lake districts, i.e., areas of young glacial relief associated with Pleistocene glaciations. The lake districts found in the north are characterized by a landscape diversified by the presence of numerous glacial lakes and postglacial accumulation forms, such as moraine hills, drumlins, eskers, sanders, and kemes.
A characteristic feature of the landscape is pre-valleys, which are wide depressions of land with flat bottoms. Formed on the foreshore of the retreating glacier, they are the result of water activities from the melting glacier and the river waters flowing from the south, which merged to form huge rivers flowing in the westerly direction, in accordance with the general slope of the European continent. The mountainous belt in southern Poland is divided into two major mountain ranges: the Sudetes in the west and the Carpathians in the east [45].
Almost the entirety of the Polish Lowlands is covered by a thick layer of postglacial sediments—clays and gravels in the moraine areas and sands in the sandstone areas. The bottoms of the river valleys and their deltas are made up of alluvium, and in some areas, there are also peat deposits. In the upland belt, there are exposed solid sedimentary rocks (limestone and gypsum). Much of the Swietokrzyskie Mountains, located in the upland belt, are composed of quartzite sandstone. The Sudetes are characterized by the most diverse geological structure. In these mountains, built mainly of crystalline rocks (especially granite), there are numerous exposures of sandstone rocks and lenses of limestone. The Polish Carpathian Mountains are mostly made of soft sedimentary rocks (sandstone and shale). Against this backdrop, two lithologically distinct ranges stand out: the Pieniny Mountains, built of limestone, and Poland’s highest mountains, the Tatras, whose eastern part is built of granite, while the western part is mainly made of sedimentary rocks (limestone) [45].
The soil cover in Poland has transitional features between the soils characteristic of Western Europe and Eastern Europe. Approximately 52% of the area is occupied by flat soils and brown earth soils, while approximately 26% of the area is occupied by rusty soils and podzols formed on sandy formations. River valleys are dominated by muds (approximately 5% of the area), while organic soils (peat soils, muck soils), gley soils, and black earths can be found in wet or humid areas. Smaller areas are occupied by rendzinas and chernozems, which are formed on carbonate rocks. The initial and poorly formed soils are characteristic of mountainous areas [45].
The climate of Poland is temperate transitional and varies from oceanic in the northwest to continental in the southeast. In the higher parts of the Sudetes and the Carpathian Mountains, the climate is mountainous. Poland is characterized by warm summers, with a mean temperature of approximately 20 °C (68.0 °F) in July and moderately cold winters, averaging −1 °C (30.2 °F) in December. Precipitation is more frequent during the summer months, with the highest rainfall amount being recorded from June to September (Figure 3).
The original natural landscape of Poland was at least 90% covered by forests, which were interrupted by vast areas of swamps and marshes, which stretched especially in the valleys and pre-valleys of the rivers and on flat, congested watersheds [48]. Treeless vegetation was associated with water reservoirs (underwater, floating, and riparian vegetation), and vegetation also developed on the bottoms of wide river valleys (meadows), on salt marshes (halophilic grasslands), on mobile dunes (psammophilous grasslands), on strongly sloped, south-facing slopes with warm microclimates (xerothermic grasslands), and above the upper forest boundary in the Carpathians and Sudetes (scrub pine and alpine grasslands).
1.1.2. Anthropogenic Factors
The beginning of significant changes in Poland’s environment took place in the Neolithic (approximately 4000 years ago) [48]. Farmers and ranchers arriving from the south and west established small, spotty settlements, especially along river valleys. For many centuries, the country’s population was low, ranging from 1 person per square kilometer in the early Middle Ages to 5 people in the 10th century and 6.6 residents in the early 16th century. By the end of the 18th century, the ratio exceeded 19, and in the mid-19th century, it reached approximately 80 inhabitants per square kilometer [49]. The environmentally significant development of urban settlements started in the late 13th century. Since then, the proportion of the population living in cities has steadily increased. In the 15th century, the urban population represented approximately 15% of the total population, and in the 16th century, the proportion reached 25%. During the same period, the coal and metal ore mining industry began to develop [49].
The development of agricultural settlements affected the wilderness landscapes by transforming them into forest-field landscapes. Multispecies deciduous forests were the prime target for burning and clearing, while pine forests, as a less favorable habitat for occupation and farming, often remained undisturbed. These forests were not resistant to urban settlements and the mining industry, which caused a surge in the demand for wood. In addition, trade was developing steadily, including wood exports. By the mid-19th century, the ratio of forests to land cover declined to 22–30%, depending on the region in Poland [49].
More significant and lasting transformations, however, started in the 19th century. Since then, the vegetation has been adversely impacted, particularly by (i) forest management focused on maximizing wood production and the large-scale introduction of monocultures of pine (Pinus sylvestris L.) and, less frequently, spruce (Picea abies (L.) H. Karst.); (ii) the intensification of agriculture, the introduction of large-scale monocultures, the use of mineral fertilizers and pesticides, the mechanization of agriculture and large-scale animal husbandry; (iii) faulty water management, especially drainage irrigation, regulation and baffling of rivers; (iv) the intensive development of heavy industry, transportation, and urbanization leading to air, water, and soil pollution and the destruction of natural habitats; and (v) spatial management, which until the end of the 20th century had little regard for the principles of sustainable development [44].
2. Material and Methods
2.1. Data Sources and Database
The database prepared for the research presented here considers national, regional, and local sources. “A checklist of Vascular Plants of Poland” was the taxonomic and nomenclatural basis [50]. The primary source of information on the occurrence of native and established alien plant species was the “Distribution Atlas of the Vascular Plants in Poland” [51,52], while for ecological data, the list was provided by Zarzycki et al. [53] and the syntaxonomic guide by Matuszkiewicz [54]. The species threat assessment was based on the list by Kaźmierczak et al. [55]. Several sources have been considered while classifying the species, taking into account their origin, time of arrival in Poland and degree of establishment. Information on archeophytes was primarily from the studies by Zając [56] and Zając et al. [57], while information on neophytes came from Tokarska-Guzik [58] and Tokarska-Guzik et al. [59], and that on casual alien species came from Urbisz [60].
As a result of many years of research at more than a dozen botanical centers, a wealth of knowledge has been developed regarding the distribution, ecology, and dynamics of the species. Approximately 2278 species (including several subspecies) are shown on the raster maps [51,52]. The ecological indicator numbers have been developed for 1772 species [53]. Almost the whole native flora was analyzed in terms of the extinction and regression of the species throughout the country. Most species in the genera Alchemilla L., Hieracium L., Oenothera L., Rosa L., Rubus L., and Taraxacum F.H. Wigg. were not evaluated for this purpose [55]. The detailed characterizations of 370 taxa (mostly at the species rank) that are extinct, declining, and at a risk of extinction were included [61].
2.2. Data Analysis Methods
The analysis, which aimed to show the changes in the flora and vegetation under human influence, was based on several commonly used classifications that take into account the features of the species that comprise it.
2.2.1. Geographic-Historical Classification
In accordance with the concept of Thellung [6,8], which has been developed in Central Europe by many authors [62,63,64,65], the classification adopted here takes into account geographical, historical, and ecological criteria (Table 1).
To standardize the terminology to obtain a better understanding between researchers in different regions and subdisciplines of biology, the classification was limited to the basic groups of species, also defined in the terms proposed by Pyšek et al. [66]. The classification of native plants and neophytes was treated more extensively. In the first case, a direct reference was made to the concept of Mirek [68], while in the second case, it was made to the division proposed by Kornaś [64].
2.2.2. Classification of Extinct and Declining Species
The criteria and threat categories adopted in the analysis are generally based on the World Conservation Union guidelines (IUCN) [69]. The basis for the classification is the knowledge of the changes in the distribution of the species, the resources of their populations, and the threats to the habitats with which these plants are associated. In the case of naturally rare species, knowledge of population resources is generally sufficient to formulate a proper diagnosis of their threat. Knowledge of the dynamic trends of the more widely distributed species is not always complete, so in this case, a criterion related to the number of sites and the habitat condition is typically used.
Compared to the IUCN guidelines [69], the classification of the extinct species has been simplified here by including them in only one group. The extinct category (EX) includes the taxa that are completely extinct (Strictly extinct according to the IUCN), extinct in the wild in their natural range (Extinct in the Wild—EW), regionally extinct (RE), and regionally extinct in the wild in their natural range (REW). The group of high-risk taxa (Threatened taxa) includes threatened species in the following decreasing order, i.e., Critically Endangered (CR), Endangered (EN), and Vulnerable (VU). There is also a category of Near Threatened (NT) and a group of threatened species that are difficult to assign a category to due to data deficiency (DD). The other species, whose dynamic trends were analyzed, were included in the category not currently at risk of extinction (NotT—Not threatened). Some of these may belong to the category of taxa of the lowest concern (LC), while others may show expansive tendencies.
2.2.3. Synecological Classification
The human impact on vegetation was analyzed at the level of synecological groups, representing the full diversity of plant communities in Poland, which are identified according to the Braun–Blanquet methodology [70]. The synecological groups were formed by combining phytosociological classes, covering plant communities with similar habitat requirements (Table 2). This made it possible to assign most species to one of the synecological groups and subject them to quantitative analysis. In the case of native species with a wider ecological scale, they were classified into the group in which they could reach their optimum occurrence. The neophytes were included in the community groups in which the species achieved the highest naturalization degree. The names of the syntaxonomic units are given according to Matuszkiewicz [54].
2.3. Methods of Developing and Presenting the Results
The assessment of the anthropogenic changes to the flora used both the absolute values and the percentage of species, representing the main geographic-historical groups and the threat categories. These values were calculated for all flora and in reference to the synecological groups. The relative proportion of species from different geographic-historical groups has been tested in studies on urban flora types in Poland [71,72,73]. The following indicators and formulas were used in this study:
- SynI (%)—synanthropization index of flora: the percentage of native species found in habitats created or disturbed by humans (apophytes) and alien species (anthropophytes) in the flora.
- ApI (%)—apophytization index of flora: the percentage of apophytes in the flora.
- AnI (%)—anthropophytization index of flora: the percentage of alien species (anthropophytes) in the flora.
- ArI (%)—archaeophytization index of flora: the percentage of archaeophytes in the flora.
- NeI (%)—neophytization index of flora: the percentage of neophytes in the flora.
- ExI (%)—extinction index of flora: the percentage of extinct and endangered species in the flora.
Due to the minor importance of casual alien species, the indicator values were mainly calculated and reported for the permanent part of the flora, which comprised the native and established alien species.
All statistical analyses and graphs were performed using Microsoft Excel spreadsheet version 6.0 for Windows.
3. Results
3.1. Geographic-Historical Structure of the Flora
According to current knowledge, Polish flora contains approximately 3614 taxa (species and subspecies), of which 73% are native and 27% are alien (Figure 4). It should be noted that although the flora of Poland seems to be well recognized, species are still being discovered. In the case of native plants, particularly significant progress should be noted for difficult and critical genera as Alchemilla, Hieracium, Oenothera, Rosa, Rubus, and Taraxacum. This group now includes approximately 650 species, which are still less studied in detail in terms of distribution and ecology. In the case of alien plants, the discovery of new species is the result of their spread and prevalence in Poland.
Approximately 11.5% are foreign plants that have not yet been established. This means that the relatively permanent flora of Poland consists of 3199 species. The vast majority of native plants (76.4%) persist exclusively in natural and seminatural habitats. A minority of native flora (23.6%) has adapted to varying degrees to man-made conditions in anthropogenic habitats. Non-synanthropic native species currently comprise 53.6% of the flora, while human-associated species (apophytes) account for 19.5% of the flora. Among the established alien taxa, neophytes comprise 11% of the flora and far outweigh archaeophytes, which comprise 4.5%.
The real importance of species representing the distinguished geographic-historical groups is their frequency in Poland. There are two distinct patterns of frequency for native plants. The vast majority of species that are limited to natural habitats (including seminatural habitats) are very rare or rare (Figure 5). The apophytes, in contrast, are common or found at a large number of sites throughout the country. There are also similar differences for permanently established alien species, with archaeophytes in this case concentrated mainly in the highest frequency classes, while very rare and rare species predominate among neophytes.
3.2. Species Decline under Human Impact
The species extinction information will be presented here with reference to the well-recognized part of the native flora (1980 species) and archaeophytes (161 species). According to a recent assessment [55], which has been slightly revised here, 764 species were classified as extinct and endangered in Poland, accounting for 35.7% of the flora considered in the study, which included 2141 species (Figure 6). Extinct species accounted for 2.2% of the flora and together with critically endangered species comprised 8%. The number of species in the next three categories was 7.1% (Endangered), 9.5% (Vulnerable) and 7.2% (Near Threatened). The threat could not be accurately determined for 3.9% of species.
The number of native plants (704 species) and the number of archaeophytes (60 species) are both decreasing under human influence. The quantitative distribution of both groups in each risk category is also similar (Figure 7).
3.3. The Species Composition of Plant Communities
The aim of the analysis was to examine the impact of species extinction and geographic-historical changes in flora on vegetation. The study included 2352 native and established alien species whose characteristics are sufficiently known to determine their synecological status at the vegetation classification level adopted in this study (Table 2). This is not always possible for recently discovered native species from critical taxonomic groups or for neophytes, even if they have already taken a permanent place in the flora.
The identified synecological groups (G) varied in terms of species richness (Figure 8). Specifically, 21.7% of the flora is made up of forest communities, including the richest zonal deciduous forests and thickets (G2 15.1%), coniferous forests (G3 3.4%), and out-of-zone deciduous forests and thickets (G1 3.2%). An even larger group of species comprise the meadow-grassland formation (28.3%), including thermophilous grasslands and ecotones (G4 16.1%), acidophilous grasslands on sands (G5 3.5%) and meadows on mineral substrate (G6 8.7%). Approximately 17.0% of the flora is composed of communities associated with water and wetland habitats. The distribution of species across the five synecological groups is fairly even in this case: aquatic communities (G11 4.5%), marshes and bogs (G9 4.4%), vegetation of muddy shores of inland waters (G10 3.3%), rushes (G7 2.7%), and coastal and inland saltmarsh communities (G8 2.1%). The mountain communities, concentrated in the southern part of the country, comprise approximately 14.7% of the flora. The richest species are high mountain grasslands and mountainous tall herbs (G14 9.3%). The proportion of species forming the vegetation of rock crevices and moving screes (G13) is 3.4%, while species associated with springs and snow bed vegetation (G11) comprise 2.1%. The overview of vegetation species richness is complemented by plants forming nitrophilous communities on forest leeks and edges (G15 4.3%) and by those adapted to ruderal and segetal habitats (G16 14.1%).
All synecological groups comprising the vegetation of Poland show traces of human influences, which are expressed in the varying participation of apophytes and alien species as well as the degrees of species threat (Table 3). The variations in the quantitative indices that describe this type of change are very wide. The synanthropization indices (SynI) vary from 1.4% in G14 to 100% in G16, with an average value of 36.1%. Native species capable of occupying anthropogenic habitats are included in each group. The indices of the apophyte share (ApI) range from 1.4% in G14 to 64.0% in G1, and the average value is 25.4%. The two groups (G13 and G14) are still free of alien species. Neophytes occur with varying abundance in the remaining groups, and additionally archaeophytes occur in groups G15 and G16. The index of the alien species share (AnI) varies from zero in G13 and G14 to 75.9% in G16, with an average value of 10.7%. It is mainly influenced by species associated with two synecological groups, i.e., G15 and G16.
The extent of the geographic-historical structure of the vegetation is more fully illustrated by the sequence of synecological groups ordered by the share of alien species (Figure 9). In the series, mountain communities free of alien species (G13 and G14) can be distinguished. They are mainly composed of native plants sensitive to human pressure. Only a few of them show the ability to occupy secondary habitats, achieving apophytic status. Similar in type are the G12, G8, and G5 groups, with only single neophytes being associated with them. In the last two, a marked increase in the participation of apophytes should be noted. In successive groups, the number of neophytes gradually increased from 4 in G11, G9, and G1 to 10 in G6. The fraction of apophytes also increased, particularly in groups G1, G10, G4, and G6. The series is closed by the three groups with the highest share of alien species (G2, G15, and G16). The species-rich G2 group is notable because of its significant proportion of native plants strongly associated with zonal deciduous forests and thickets, and these species cannot occupy anthropogenic habitats. A completely separate, specific character is found for the G16 group, which combines three categories of plants associated with human activity: apophytes, archaeophytes, and neophytes. Native species are the minority and are represented only by plants adapted to secondary habitats. Among alien species, archeophytes outnumber neophytes.
The threat to the flora varies greatly among the distinguished synecological groups (Table 3). The highest proportion of threatened species is found in the group of coastal and inland saltmarsh communities (G8 79.6%) and marshes and bogs (G9 69.6%). Over half of the species associated with freshwater communities (G11 53.9%) and communities occupying muddy shores of inland waters (G10 51.4%) are threatened. A slightly lower level of threat applies to species that form dry and thermophilic grasslands and ecotones (G4 49.1%). The range of threatened species is 31.3–40.0% and includes three groups of mountain communities (G12, G13, G14) as well as coniferous forests (G3) and acidophilous heaths and poor grasslands (G5). A relatively lower degree of threat (20–26%) is found in the groups of species associated with meadows on mineral substrates (G6), rushes (G7) and deciduous forests and thickets (G2). A similar level is represented by a group of ruderal and segetal communities (G16). In this case, the fraction of endangered species is formed exclusively by archaeophytes. Significantly fewer declining species are associated with nitrophilous communities developed on forest leeks and edges (G15) and azonal deciduous forests (G1).
The participation of species from each synecological group in the threat to the flora of Poland is presented in a sequence in which they are ordered by a decreasing extinction coefficient (Figure 10). This overview shows that extinct species are represented in most synecological groups, with the exception of groups G1, G3, and G7. These taxa are most abundant in groups G6, G8, G11, and G14. The Red List of the vascular flora of Poland is strongly dominated by representatives of thermophilous grasslands. The share of the G4 group is approximately 23.7%. It is followed by groups G14, G9, and G2, though they have a significantly smaller participation.
3.4. Process of Human-Influenced Extinction and Expansion of Species
3.4.1. Natural and Anthropogenic Extinction Factors
The causes of plant decline were analyzed in a group of 370 species that were best identified for anthropogenic threat in Poland [61]. Based on the interpretation of the records of the researchers visiting the sites of declining plants, a list of factors responsible for the extinction of each species was generated, and the areas of human activity to which they are linked were identified. The list of factors contains 768 items, including 98 describing the natural determinants of extinction and nearly 640 anthropogenic factors, which were assigned to nine areas of human activity (Figure 11).
Among the natural determinants, rarity and low population size are mentioned most. In extreme cases, there are single sites with a few individuals each. The authors of the studies also note the location of the sites outside or at the limits of the range, the insular nature of these sites, the very strict habitat requirements, the tendency to hybridize and the presence of genetic erosion, the vulnerability of parasite–host systems, the low resistance to various animal influences (e.g., stem gnawing), and random phenomena. Species exposed to extinction due to natural determinants are particularly abundant in montane communities (G12, G13, G14) and in thermophilous grasslands (G4) (Figure 12).
The list of anthropogenic threats to species opens with plant collecting and hoarding. Wild plants, including very rare species, continue to attract interest due to their useful nature (medicinal, ornamental, etc.). They continue to be collected and transplanted to private gardens. Unfortunately, there are known examples of plant populations being threatened by the overcollection of specimens for scientific purposes. This direct threat factor was mainly identified for upland (G13 and G14), forest (G2 and G3), and grassland (G4) species.
Significant threats to declining plants come from forest management. Unfortunately, these threats are not limited to forest communities, especially groups G2 and G3; they also affect grasslands (G4 and G5) and mountain communities (G13 and G14). Threat factors generated by forestry impact populations of endangered species directly or indirectly by disturbing the phytocoenosis structure or transforming habitat conditions. Populations of some forest species are threatened directly during various silvicultural procedures. The authors of threat assessments identify, among other things, the following activities: planting of conifers, primarily Pinus sylvestris L., in deciduous forest habitats, introduction of alien species (e.g., Padus serotina (Ehrh.) Borkh.), clear-cutting forestry, damage during forestry activities, and the elimination of traditional forms of forest use such as grazing, litter raking, and brushwood collection. Threats to forest species with specific ecological requirements are also associated with the use of chemicals for pest control, mineral fertilizers, progressive eutrophication of habitats, and drainage, which is part of the separately discussed water management. These factors have the effect of activating such lines of succession that lead to changes in light, microclimatic conditions, and competition, ultimately eliminating the most vulnerable ecosystem components. The effects of forest management on grassland species are primarily associated with the afforestation of open areas, while the impact on mountain communities is associated with the planting of Pinus mugo Turra.
The greatest threat to retreating plants is agriculture, which, in addition to arable farming, includes pastoralism and fish farming. Approximately 33.3% of the factors responsible for the extinction of the analyzed group of species are related to this form of human activity. They negatively affected species representing 12 synecological groups. The most affected species are those associated with field habitats (G16), meadows (G6), grasslands (G4 and G5), and aquatic communities (G11). A particularly interesting case concerns weeds in arable fields that are archaeophytes, whose presence in the flora of Poland is the result of human activity. After many centuries of functioning in agrocoenoses, some of these species are becoming extinct due to agrotechnical changes and agricultural intensification. Factors such as the reduction of certain crops (e.g., flax), seed purification, the introduction of prolific and fast-growing cereal varieties, ploughing immediately after harvesting, and the use of plant protection products, herbicides, and mineral fertilizers have a strong influence the extinction of field archaeophytes.
For other reasons, meadow and pasture species (G6) and grassland species (G4, G5, and G14) are at risk of extinction. A key factor in these cases is the inappropriate method and intensity of their use. The meadow species are threatened by both overmowing and abandonment. In the case of grasslands, traditional uses such as mowing, grazing, or burning have been reduced or completely eliminated. A longer break in the established use of meadows and grasslands over the years triggers a succession involving tall perennials, shrubs, and trees, resulting in progressive changes in habitat conditions that determine the occurrence of many rare species. In the lowlands, both native species (e.g., Bromus inermis Leyss., Calamagrostis epigejos (L.) Roth, Elymus repens (L.) Gould, Prunus spinosa L., Rubus caesius L., Cornus sanguinea L., Frangula alnus Mill.) and non-native species (e.g., Robinia pseudacacia L.) are involved in this process. Native trees Sorbus aucuparia L. em. Hedl. and Alnus viridis (Chaix) DC. in Lam. & DC. participate in the overgrowth of mountain grasslands. Succession causes an increase in competition and a change in abiotic conditions, including increased shading and nutrient enrichment of the ground. Eutrophication is also caused by the runoff of mineral fertilizers from the surrounding agricultural fields. In the same way, chemical plant protection chemicals penetrate meadow and grassland habitats.
Eutrophication, humification, and excessive acidification are the main reasons for the decline in aquatic (SG11) and wetland habitat species (G7, G8, G9, G10) caused by agriculture. In the case of vegetation of muddy shores of inland waters (G10), factors related to fisheries management also have a negative effect.
Species from the listed plant communities and the meadow plants (G6) are most vulnerable to factors generated directly by water management, which aim to create conditions for the use of inland surface and groundwater resources in various areas of human activity. The effect of amelioration is to drain wetlands, peat bogs, saltmarshes, and swampy forests. This leads to a succession involving expansive perennials, shrubs of the genus Salix L., and trees of the genera Betula L. and Alnus Mill. Some plant species are also negatively affected by the long-term stagnation of water. The important water management tools include negative effects on sensitive plant species during the regulation of rivers and streams, the construction of dams and dikes and the installation of various types of hydraulic engineering equipment. These activities cause the direct destruction of plant populations and contribute to the permanent transformation of rush communities (G7) and muddy shore vegetation (G10).
Two further areas of human activity influence species populations and habitats that survive in landscapes previously altered by agriculture and water management. The development of settlements, urban extensions, and various types of investments are very often associated with the devastation of vegetation and fragmentation of habitats leading to their complete transformation. Land development for housing, transport, and industrial purposes ultimately leads to the elimination of plant populations that are highly sensitive to human pressure as well as species that are adapted to extensive forms of human activity. Habitat ruderalization opens up space for a relatively small group of native species (apophytes) and alien plants. The towns and industrial centers generate pollution, which contributes to the eutrophication, humification, and acidification of the water and the terminal elimination of many species comprising freshwater vegetation (G11). The expansion of urban and settlement areas, as well as road and industrial developments, is based on raw materials extracted close to their location or from a considerable distance in places with rich naturally rare plant species. Peat exploitation threatens species found in bogs and mires (G9), gravel extraction adversely affects grasslands (G4), and quarry enlargement has resulted in the loss of rare forest species (G2).
A growing factor on the list of human activities that threaten flora and vegetation is tourism, recreation, and sport. The effects of this type of activity are visible in most plant communities but are especially apparent in the vegetation communities of rock crevices and moving scree (G13) and alpine grasslands (G14). It is worth noting that mountain tourism is a direct and greatest threat to these communities among all forms of human pressure. This also applies to advanced tourism and sports, including mountaineering and skiing, the negative traces of which have already been identified in the Tatras. In the lowlands, the destructive impacts on coastal halophytic communities (G8) and freshwater vegetation (G11) are most evident. The following factors contribute to this negative impact: the establishment of recreational plots on previously drained salt marshes, the removal of organic debris dumped by waves on the seashore that harbors annual halophytes, the destruction of coastal dunes, the development of lake shores, the destruction of the littoral zone, and eutrophication.
At least part of the responsibility for the decline of the species analyzed here must be attributed to global, or supra-regional, environmental changes. Usually, however, it is difficult to separate these from the local causes already described because they most often operate together. Nevertheless, it is worth noting at least two supra-regional factors that have a significant impact on plant populations most sensitive to anthropogenic change. First, the prolonged periods of drought resulting from climate change have been identified as an important threat to species in the spring and snow plant communities (G12). Second, atmospheric pollutant emissions are included among the causes of eutrophication of xerothermic grasslands (G4) and subsequent succession.
3.4.2. The Process of Human-Induced Spread of Native Species
The ability of native plants to break through ecological barriers and occupy man-made habitats (i.e., apophytism) shows a wide range of variability that allows species to be classified into several groups (Figure 4; Table 4A). The first, and by far the most abundant group, includes species never recorded in alternate habitats. Native non-synanthropic species (nsNS) are represented in almost all synecological groups (except G16), frequency classes, and threat categories. The second group is made up of native species that ephemerally appear in secondary habitats (Table 4B). This includes species that are rare by nature and declining in primary stands. However, such limited adaptive ability may not be sufficient to persist in the long term. The third group is made up of apophytes proper, i.e., species that are native to natural and seminatural communities and, at the same time, permanently established in man-made habitats (Table 4C).
Most of these are currently widespread or common species, but endangered species are also found in this group. Secondary habitats very often contain species derived from moist and wet forests and mesotrophic deciduous forests, as well as forest clearings and coppice. Plants from various types of grasslands and meadows and thermophilous scrubs play a very important role in the human-accompanying communities. Saltmarshes, rush species, and plants of muddy banks of inland waters often spread from moist and wet habitats. Artificial bodies of water created for utility purposes are often overgrown with plants brought in from natural bodies of water. A particularly interesting apophytic pattern is represented by species that retreat from natural communities while simultaneously increasing their range in man-made habitats. These include halophytes, which decline in natural salt marshes and yet spread to secondary saline sites. With this ability, species have a good chance of surviving and even expanding their range. The fourth group of apophytes includes species whose native status is unlikely to be in doubt, although it is currently difficult to identify the natural community that was the origin (Table 4D).
3.4.3. Establishment and Naturalization of Alien Species
Several steps can be distinguished in the process of establishment and naturalization of geographically alien species, and these steps dictate how they are divided into groups.
The first group includes casual alien species, i.e., species introduced accidentally usually by transport (ephemerophytes) or alien plants escaping from cultivation sites (ergasiophytes) (Table 5A). Casual alien species form the largest group of non-native species (Figure 4), and although their importance in the vegetation structure is negligible, they deserve attention because they represent the plants that may potentially become established in the future. It should be noted that it is not possible to estimate casual alien species appearing in the distant past. It is therefore not possible to indicate how large the group of species from which the oldest non-native species originated from.
The second group of alien species includes archaeophytes, whose occurrence is in principle restricted to man-made habitats. Only a few of them are recorded in near seminatural communities, e.g., on the edges of forest and scrub communities (Anthriscus caucalis M. Bieb., A. cerefolium (L.) Hoffm. and Parietaria officinalis L.). Archaeophytes are important in field weed communities and ruderal habitats (Table 5B1). They are dominated by frequent, very frequent, and common species (Figure 5). The most specialized of these species are declining (Figure 7), primarily due to the influence of agricultural intensification (Figure 12).
The direction and rate of change over the last 200 years is mostly determined by neophytes, which are found in almost all plant communities with the exception of two synecological groups related to montane vegetation (Table 3). The establishment of neophytes in natural communities is usually the culmination of a process in which several stages can be distinguished. The process begins with the accidental importation of an alien species or the ‘escape’ of an alien species from a cultivation site. The potential for casual alien species is increasing due to the rapid development of the road network and increased transport, as well as the ever-expanding range of plants grown for ornamental purposes (currently more than 400 species). As a result, the number of alien species established in Poland in recent years has increased (Table 5B2). Most of the species spreading in this way occupy only anthropogenic habitats and reach epecophyte status. There are currently approximately 275 species in this phase of expansion. In man-controlled habitats, epecophytes do not face much competition, but together with apophytes and archaeophytes, they build communities of ruderal and segetal weeds. Similar to archeophytes, specialists associated with cultivated fields or ruderal communities can be observed in epecophytes.
The ecological barriers separating anthropogenic habitats from semi-natural and natural communities are much more difficult to break through. The analysis shows that 123 neophytes, including 100 haemiagriophytes and 23 holoagriophytes, have overcome ecological barriers in Poland (Table 5B2). In various types of forest (G1, G2, and G3), 45 species have established themselves; in grasslands (G4, G5) and meadows (G6), 20 species have been established; and in waters (G11), waterbanks (G7, G8, G10), and peatlands (G9), 22 species have established. The other agriophytes are associated with natural and seminatural nitrophilous communities developed in fresh habitats and occur in moist or wet ecotones at the transition zone between woody and herbaceous communities (G15).
4. Discussion
4.1. Main Directions of Anthropogenic Flora Changes
The human-influenced changes in the flora and vegetation of Central Europe are a historical process rooted in the beginnings of agricultural settlement, which definitely accelerated in the 19th century with the development of industry, transport, and urbanization [33,34,36,38,74,75]. The traces of human development have been imprinted in the species composition, structure, and dynamics of modern flora and vegetation throughout the region to varying degrees of intensity. The results of studies carried out in Poland show that centuries of human activity and external influences, including global effects, have had very significant consequences for plant diversity. They confirm the main theses formulated in the 1960s and are still in use concerning the directions of changes in the country’s flora and vegetation [33,34,75]; however, for the first time, they document, in the form of quantitative indicators, the most important aspects of plant cover synanthropization. In this way, both the species receding and expansion phenomena and the causes and consequences of both at the plant community level are presented.
In assessing the present state of Polish flora and vegetation, it is necessary to remember that in the Neolithic, more than 90% of Central Europe was covered by forests [76]. Currently, approximately 30% of Poland’s area is covered by forests, which mainly perform economic functions [49]. Primeval forests occupy a small area. Flora synanthropization began around mid-forest settlements with the selection of native plants [77,78]. Its first effect was the separation of native species adapted to transformed or man-made habitats (apophytes). The spread of native plants into anthropogenic habitats, known as flora apophytization [36,71,79,80,81,82,83,84], consists of breaking down barriers separating natural from non-natural ecosystems and is therefore also called ecological expansion [85].
The first wave of the immigration of alien species, the end of which was close to the symbolic date of the discovery of America (1492), was associated with the development of agricultural settlements. The occurrence of archaeophytes together with apophytes in medieval Poland is confirmed by numerous archaeobotanical discoveries [86,87,88].
Following the hypothesis of Fukarek [75], confirmed by studies in Poland [33,34] in the 18th century, the flora of Central Europe was successively enriched by newcomers. The wave of the inflow of Neophytes into the Polish area accelerated in the 19th century [58,89]. The phenomenon of neophytization of flora and vegetation, in contrast to the influx of archaeophytes, is monitored in Central Europe on an ongoing basis. It is a multistage process including an introduction phase and several stages of naturalization [33,34,36,75,79,80,90,91,92]. The crucial moment is the crossing of the species’ natural range limit; therefore, this process is referred to as geographical expansion [85]. According to Udvardy’s [93] concept, which was tested in our research, the already mentioned geographical expansion and ecological expansion are two phases of the same expansion process. Fukarek [75] notes that since the 19th century, the increase in the number of alien species has not compensated for the loss of plant diversity resulting from the decline of native species and archaeophytes under the influence of the ‘industrial revolution’. This thesis is true in light of the research carried out; however, it is not true in terms of the numerical statement with regard to the ecological effects of extinction and expansion. The alien species not only fail to fulfil the role of native species but are also one of the factors contributing to their extinction.
4.2. Native Species Adaptation to Man-Made Habitats
Our study shows that the native flora of Poland includes 2639 taxa (species and subspecies), and this number is mainly due to the inclusion of new discoveries and captures in several critical taxonomic groups. In comparison with other Central European countries, the Polish native flora is richer than that in the Czech Republic (2401 taxa) [94] and poorer than the floras of Germany (2743) [95] and Slovakia (3344) [96].
Approximately 35.4% (n = 1992) of the native species examined have adapted to habitats under strong human pressure. With the lack of comparative data, it is difficult to compare these indicators to other European countries. However, it is worth noting that the proportion of apophytes changes directionally in the comparison of flora of areas with different intensities of land use. For example, in protected areas in Poland, in Słowiński National Park [97] and Wielkopolski National Park [98], the proportion is 28.2% and 32.5%, respectively. In the large cities of Poland, such as Poznań [71] and Warsaw [99], it reaches values of 42.0% and 49.5%, respectively. In an agricultural region such as the Gniezno Lake District [100], the proportion of apophytes in the native flora is 47.0%. The highest values are reached in industrial areas; for example, in Silesia [101], it is as high as 61%.
4.3. Introduction and Naturalization of Alien Species
The human impact on the flora is most evident through the inflow of geographically foreign plants. The modern European flora includes 2843 alien species from outside the continent and as many as 5789 species recognized as alien in at least some countries [102]. There are currently 975 alien species identified in Poland. Previously, 1017 [58] species were reported, which is mainly due to the changing interpretation of the status of cultivation-derived neophytes. Changes in the numbers of alien species in the floras of European countries are quite common, and these changes are due not only to the increasing expansion of alien species but also to advances in the knowledge of the expansion process. The comparisons are also hampered because we do not always have complete data, and different classification systems are still in use, despite attempts to standardize them [66,103].
In presenting Polish flora in comparison to other European countries, we will refer to the following sources: the Czech Republic [94], Germany [95], Slovakia [96], Ukraine [104], Estonia, Lithuania, Latvia, Slovenia, Hungary, Austria, Switzerland, and other countries [102]. In the 12 countries listed above, there are an average of 831.3 alien species. The fewest anthropophytes were reported in Switzerland (313 species) and Estonia (416), while the most were reported in Austria (1086) and the Czech Republic (1350). A lower number of alien species than that in Poland was recorded in other Central and Central-Eastern European countries, i.e., Germany and Slovakia (916 species) and Ukraine (830 species). These changes are more fully illustrated by the percentage index of the proportion of anthropophytes (AnI), which is 27.0% for the total flora of Poland. This index has a higher value in the Czech Republic and a lower value in Germany (25.0%), Slovakia (21.5%), and Ukraine (13.8%). The share of alien species in the flora of Poland is similar to that in some countries outside Central Europe, e.g., Denmark, Spain, and Finland, where the number of anthropophytes ranges from 900 to 1000 species. In contrast, Poland has fewer alien species than Wales, Austria, Scotland, Sweden, and France, where this parameter remains in the range of 1000–1500 species, and in England and Belgium, where it significantly exceeds 1500 species [102]. The number of established taxa places Poland second in the group of Central European countries (560 species). There are more in Germany (709) and fewer in the Czech Republic (490) and Slovakia (388). The percentage indices of established anthropophytes in the casual alien species-free flora in these countries are as follows: Germany (20.5%), Poland (17.5%), the Czech Republic (17.0%), and Slovakia (10.4%). The number of naturalized anthropophytes in the other countries from the analyzed group is considerably lower and is as follows: Slovenia (330 species), Latvia (303), Austria (276), Lithuania (258), Switzerland (175), Hungary (145), and Estonia (125). In addition, Poland has a similar number of established alien species as that found in Norway (576), while in Spain, it is slightly lower (495) [102].
The centuries-long influence of agriculture was imprinted in the presence of alien species introduced before the end of the 15th century. A detailed study of these plants in Poland was carried out and resulted in the identification of 161 archaeophytes [56,57,105,106,107,108]. This is a comparable number to the Ukrainian flora (149 species) and significantly lower than the number of archaeophytes found in Germany (258), Slovakia (282) and the Czech Republic (309). The percentage of archaeophytes in the floras of these countries devoid of casual alien species is as follows: Ukraine (2.5%), Poland (5.0%), Germany (7.5%), Slovakia (7.6%), and the Czech Republic (10.7%). The explanation for such significant differences requires a detailed analysis, but it seems that they are more likely to occur due to the differences in the assessment of the origin of species than from the environmental conditions.
The civilization changes initiated in the 19th century activated another wave of alien plants, a considerable number of which (399 species) have found a permanent place in the flora of Poland. The number of established neophytes in Poland is comparable to those in countries such as Germany (424), Italy (440), Belgium (447) and Ukraine (454). It must be highlighted that the neophyte’s contribution has been updated several times, due to both the ongoing introduction of these species and the progress in recognizing this phenomenon. The first lists were published in the 1970s [109], a comprehensive study was published in 2005 [58], and another compilation was published in 2012 [59].
The introduction of alien species into Poland is continuing, and their potential to establish increases continuously. Most of these species are casual alien plants, of which there are currently at least 415 species. This is lower than the floras of Germany (207), Slovakia (528), Hungary (566), Austria (810), and the Czech Republic (891) but higher than those of the United Kingdom (395), Spain (362), and Lithuania (360). The relatively low number of anthropophytes in the German flora is probably due to a more restrictive approach in which escaped plants are included in the spontaneous flora.
4.4. Human-Induced Species Decline
The process of decline in the flora of Poland is taking place in parallel with the adaptation of native plants to anthropogenic habitats and the introduction and naturalization of alien species. Due to human influence, not only native species but also archaeophytes are disappearing. The study showed that 35.6% (n = 1980) of the analyzed part of the native flora, i.e., 29.0% (n = 2639) of the total native flora, was in various stages of decline. However, it is important to add that the Polish Red List of Ferns and Flowering Plants does not yet include species potentially belonging to the least concern (LC) category. In Poland, the number of receding species increased from approximately 500 to 765 taxa (including archaeophytes) between 2006 and 2016, in which successive editions of the Red List were published [55]. In the author’s view of the latest study, this is less due to the increasing threat to species than to an increase in knowledge of plant declines.
The comparison of the threat to the flora of Poland to that in other European countries is limited, which is caused, among others, by (i) the varying state of research development, (ii) different publication times, (iii) incomplete data, and (iv) differences in the classification criteria for receding species. Some indicators are worth noting, despite these caveats, because they illustrate differences in the extent of flora synanthropization. We will consider data from Slovakia [110], the Czech Republic [111], Italy [112], Spain [113], England, and Wales [114], mainly concerning the extinct (Ex, EW, RE) and threatened (CR, EN, and VU) categories, which seem to be the most comparable ones.
On average, 57 species were categorized as extinct, the least in Italy (13), and the most were found in the Czech Republic (156, including 53 missing taxa and 29 uncertain cases). The number of species considered extinct in Poland (44 native species and 4 archaeophytes) is higher than the number of species extinct in Wales (38), England (36), and Spain (25) but almost half of that in Slovakia (83). On average, 659 species were classified as threatened in the countries compared, e.g., Wales (227) and England (370), the most in Spain (1196) and again in the Czech Republic (1184). The number of endangered taxa in Poland (442 native species and 38 archaeophytes) is close to that in Slovakia (527) and comparable to that in Italy (631).
The countries analyzed differed fundamentally in their overall flora species richness, so it is worth comparing the percentage extinction rates in the countries for which the total number of species is given in cited sources. The proportion of extinct and endangered species in the flora of Poland is 20% (n = 2639) and is significantly higher than that in Italy (7.9%; n = 8195), higher than that in Slovakia (16.9%; n = 3619) but almost half that in the Czech Republic (46.1%; n = 2905). From the data provided above, it can be seen that plants receding in Poland are at an average European level and are in many respects similar to the situation in Slovakia.
A species threat categorization according to IUCN standards primarily has conservation objectives. The diagnoses formulated on the basis of expert analyses, not devoid of subjectivity, serve this purpose. The Red Lists, and even more so the Red Books, make a major contribution to the knowledge of flora synanthropization, in particular the biology and history of disappearing species, the reasons for their decline, and threatened plant communities and habitats.
4.5. Human-Influenced Vegetation Changes
The taxonomical composition and structure of plant communities depend on abiotic and biotic factors, among which the pool of species forming a given area’s flora plays an important role. The analyses of the 16 plant community groups showed that they differed significantly in terms of the proportion of native and non-native plants, as well as in the proportion of declining species. The naturalization process of non-native species was found in 14 plant community groups. Alien species are most abundant in ruderal and segetal habitats and on leeks and woodland edges and form entirely human-dependent communities with apophytes. Neophytes are part of zonal and azonal forest communities, grasslands and various grasslands, all types of aquatic communities, and wetland habitats. To date, they have not established themselves only in high mountain grasslands, mountain herbs, rock crevice communities, and moving screes. The extinction process affects all groups of plant communities, including human-made communities. The coastal and inland halophilic communities, peat bogs, and marshes are most affected.
The causes of human-influenced flora and vegetation changes in Central Europe are already well identified, in terms of both the process of species decline and the expansion of alien taxa. The expansion of alien species is contributed mainly to the development of agriculture, trade, transport and urbanization [21,31,36,58,115,116], which lead to fundamental changes in land use [42]. The list of factors responsible for species extinction includes collecting and hoarding, forestry, agriculture, water management, settlement and urbanization, investment activities, mining, tourism, recreation, and sport, and global environmental changes, primarily climate warming [36,38,74,75,112,117]. Our research has confirmed the previously identified reasons for the decline of species, and in principle, it does not contradict their prioritization, according to which the greatest influence is attributed to agriculture [20,117]. The results clearly indicate, however, that the importance of anthropogenic factors varies according to plant community types, habitats, and time [118]. For example, a significant influence of the oldest factor, namely, collecting, was found in high-mountain and mountain communities, xerothermic grasslands and forest communities. In contrast, agriculture plays a key role in the extinction of species in all communities, with the exception of rocky vegetation. The species composition and the structure of mountain communities, freshwater vegetation, and halophilic communities are increasingly influenced by tourism, recreation, and sport. In the high mountains, where the direct human impact on vegetation is quite limited, the negative impact of global climate change has been identified.
We have shown at the national scale that, as at the global scale [40], extinction of native species and naturalization of non-native species change the shape of native plant communities. This leads to a profound homogenization of biodiversity [40], as already pointed out when defining the process of synanthropization of flora and vegetation [37,39].
Funding
This work was supported by the Faculty of Biology of the Adam Mickiewicz University in Poznań under the Research Subvention (Number: 4102000000-604-506000-BN002023) and the European Union through the European Regional Development Fund under the Operational Program Digital Poland (OP PC), Priority axis: II E—administration and open government; Action: 2.3 Digital accessibility and usability of public sector information; Submeasure: 2.3.1 Digital access to public sector information from administrative sources and science resources (Grant numbers POPC.02.03.01-00-0043/18-05).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The datasets generated and/or analyzed during the current research are available from the author at the Department of Systematic and Environmental Botany at the Faculty of Biology of Adam Mickiewicz University, Poznań, [email protected].
Conflicts of Interest
The author declares no conflict of interest.
References
- Roberts, N. The Holocene. An Environmental History, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2014; 372p. [Google Scholar]
- Steffen, W.; Crutzen, P.J.; McNeill, J.R. The Anthropocene: Are Humans Now Overwhelming the Great Forces of Nature? In Environment and Society: A Reader; Schlottmann, C., Jamieson, D., Colin Jerolmack, C., Anne Rademacher, A., Eds.; New York University Press: New York, NY, USA, 2017; pp. 12–31. [Google Scholar] [CrossRef]
- Behre, K.E. The role of man in European vegetation history. In Vegetation History. Handbook of Vegetation Science; Huntley, B., Webb, T., Eds.; Springer: Dordrecht, The Netherlands, 1998; Volume 7, pp. 633–672. [Google Scholar]
- Watson, H.C. Cybelle Brittanica: Or British Plants and Their Geographical Relations, I; Longmans: London, UK, 1847; 560p. [Google Scholar]
- Candolle, M.A. De, Géographie Botanique Raisonnée ou Exposition des Faits Principaux et des lois Concentrant La Distribution Geographieque des Plantes de l’epoque Actuelle; Librarie de Victor Masson: Paris, France; Geneva, Switzerland, 1855; 1366p. [Google Scholar]
- Thellung, A. Einteilung der Ruderal und Adventivflora in genetische Gruppen. In Die Flora des Kanton Zürich. I. Teil. Die Ruderal- und Adventivflora des Kanton Zürich; Naegeli, O.O., Thellung, A., Eds.; Vierteljahresschr. Naturforsch. Ges.: Zürich, Switzerland, 1905; Volume 50, pp. 232–236. [Google Scholar]
- Thellung, A. Pflanzenwanderungen unter dem Einfluss des Menschen. Englers Bot. Jb. Leipzig. 1915, 53, 37–66. [Google Scholar]
- Thellung, A. Zur Terminologie der Adventiv und Ruderalflora. Allg. Bot. Zeitschr. Karlsr. 1919, 24, 36–43. [Google Scholar]
- Thellung, A. Kulturpflanzen-Eigenschaften bei Unkräutern. Veröff. Geobot. Inst. Rübel Zürich 1925, 3, 745–762. [Google Scholar]
- Linkola, K. Studien über den Einfluss der Kultur auf die Flora in den Gegenden nördlichem Ladegasee. I. Allgemeiner Teil. Acta Soc. Faun. Flor. Fenn. 1916, 45, 1–432. [Google Scholar]
- Zerbe, S. Restoration of natural broad-leaved woodland in Central Europe on sites with coniferous forest plantations. For. Ecol. Manag. 2002, 167, 27–42. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.-L.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Tockner, K.; Stanford, J. Riverine flood plains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef]
- Nobis, A.; Żmihorski, M.; Kotowska, D. Linking the diversity of native flora to land cover heterogeneity and plant invasions in a river valley. Biol. Conserv. 2016, 203, 17–24. [Google Scholar] [CrossRef]
- Gołdyn, H. Changes in plant species diversity of aquatic ecosystems in the agricultural landscape in West Poland in the last 30 years. Biodivers. Conserv. 2010, 19, 61–80. [Google Scholar] [CrossRef]
- Olson, E.R.; Doherty, J.M. Macrophyte diversity–abundance relationship with respect to invasive and native dominants. Aquat. Bot. 2014, 119, 111–119. [Google Scholar] [CrossRef]
- Pavlů, L.; Pavlů, V.; Gaisler, J.; Hejcman, M.; Mikulka, J. Effect of long-term cutting versus abandonment on the vegetation of a mountain hay meadow (Polygono-Trisetion) in Central Europe. Flora 2011, 206, 1020–1029. [Google Scholar] [CrossRef]
- Wesche, K.; Krause, B.; Culmsee, H.; Leuschner, C. Fifty years of change in Central European grassland vegetation: Large losses in species richness and animal-pollinated plants. Biol. Conserv. 2012, 150, 76–85. [Google Scholar] [CrossRef]
- Baessler, C.; Klotz, S. Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years. Agric. Ecosyst. Environ. 2006, 115, 43–50. [Google Scholar] [CrossRef]
- Rühl, A.T.; Eckstein, R.L.; Otte, A.; Donath, T.W. Future challenge for endangered arable weed species facing global warming: Low temperature optima and narrow moisture requirements. Biol. Conserv. 2015, 182, 262–269. [Google Scholar] [CrossRef]
- Kowarik, I. On the role of alien species in urban flora and vegetation. In Plant Invasions-General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 85–103. Available online: https://link.springer.com/content/pdf/10.1007/978-0-387-73412-5.pdf#page=333 (accessed on 1 March 2023).
- Jackowiak, B. Dynamik der Gefäßpflanzenflora einer Großstadt am Beispiel von Poznan/Polen. Braun-Blanquetia 1989, 3, 89–98. [Google Scholar]
- Pyšek, P. Alien and native species in Central European urban floras: A quantitative comparison. J. Biogeogr. 1998, 25, 155–163. [Google Scholar] [CrossRef]
- Kühn, I.; Brandl, R.; Klotz, S. The flora of German cities is naturally species rich. Evol. Ecol. Res. 2004, 6, 749–764. Available online: https://www.evolutionary-ecology.com/abstracts/v06/1629.html (accessed on 1 March 2023).
- Jehlîk, V.; Dostálek, J.; Frantík, T. Alien plants in Central European river ports. NeoBiota 2019, 45, 93–115. [Google Scholar] [CrossRef]
- Jehlîk, V. Beitrag zur synanthropen (besonders Adventiv-) Flora des Hamburger Hafens. Tuexenia 1981, 1, 81–97. [Google Scholar]
- Brandes, D. Flora und Vegetation der Bahnhöfe Mitteleuropas. Phytocoenologia 1983, 11, 31–115. [Google Scholar] [CrossRef]
- Wittig, R. Ferns in a new role as a frequent constituent of railway flora in Central Europe. Flora 2002, 197, 341–350. [Google Scholar] [CrossRef]
- Wrzesień, M. Kenophytes chorologically related to the habitats of railway grounds in central eastern Poland. Biodiver. Res. Conserv. 2006, 1, 92–94. Available online: http://brc.amu.edu.pl/Kenophytes-chorologically-related-to-the-habitats-of-railway-grounds-in-central-eastern,121552,0,2.html (accessed on 1 March 2023).
- Jehlík, V.; Zaliberová, M.; Májeková, J. The influence of the Eastern migration route on the Slovak flora—A comparison after 40 years. Tuexenia 2017, 37, 313–332. [Google Scholar]
- Lippe, M.; Kowarik, I. Long-Distance Dispersal of Plants by Vehicles as a Driver of Plant Invasions. Conserv. Biol. 2007, 21, 986–996. [Google Scholar] [CrossRef]
- Kornaś, J. Influence of man and his economic activities on the vegetation of Poland. The synanthropic flora. In The Vegetation of Poland; Szafer, W., Ed.; Pergamon Press-PWN: Oxford, UK; Warszawa, Poland, 1966; pp. 93–137. [Google Scholar]
- Kornaś, J. Man’s impact upon the flora: Processes and effects. Memorab. Zool. 1982, 37, 11–30. Available online: https://rcin.org.pl/miiz/dlibra/publication/edition/38769 (accessed on 1 March 2023).
- Kornaś, J. Man’s impact upon the flora and vegetation in Central Europe. In Man’s Impact on Vegetation; Holzner, W., Werger, M.J.A., Ikusima, I., Eds.; Dr. W. Junk Publishers: The Hague, The Netherlands; Boston, MA, USA; London, UK, 1983; pp. 277–286. [Google Scholar]
- Sukopp, H. Der Einfluß des Menschen auf die Vegetation. Vegetatio 1969, 17, 360–371. [Google Scholar] [CrossRef]
- Sukopp, H. Wandel von Flora und Vegetation in Mitteleuropa unter dem Einfluß des Menschen. Ber. Ldw. 1972, 50, 112–139. [Google Scholar]
- Faliński, J.B. Synantropizacja szaty roślinnej—Próba określenia istoty procesu i głównych kierunków badań. Phytocoenosis 1972, 1, 157–170. (In Polish) [Google Scholar]
- Sukopp, H.; Trepl, L. Extinction and Naturalization of Plant Species as Related to Ecosystem Structure and Function. In Ecological Studies; Schulze, E.D., Zwolfer, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; Volume 61, pp. 245–276. Available online: https://link.springer.com/chapter/10.1007/978-3-642-71630-0_12 (accessed on 1 March 2023).
- Faliński, J.B. The interpretation of Contemporary Vegetation Transformations on the Basis of the Theories of Synanthropisation and Syndynamics. In Mechanism of Anthropogenic Changes of the Plant Cover; Jackowiak, B., Żukowski, W., Eds.; Publ. Dep. Plant Taxon. AMU: Poznań, Poland, 2000; Volume 10, pp. 9–30. [Google Scholar]
- Daru, B.H.; Davies, T.J.; Willis, C.G.; Meineke, E.K.; Ronk, A.; Zobel, M.; Pärtel, M.; Antonelli, A.; Davis, C.C. Widespread homogenization of plant communities in the Anthropocene. Nat. Commun. 2021, 12, 6983. [Google Scholar] [CrossRef]
- Bartolucci, F.; Galasso, G.; Peruzzi, L.; Conti, F. Report 2020 on plant biodiversity in Italy: Native and alien vascular flora. Nat. Hist. Sci. 2021, 8, 41–54. [Google Scholar] [CrossRef]
- Scherrer, D.; Bürgi, M.; Gessler, A.; Kessler, M.; Nobis, M.P.; Wohlgemuth, T. Abundance changes of neophytes and native species indicate a thermophilisation and eutrophisation of the Swiss flora during the 20th century. Ecol. Indic. 2022, 135, 108558. [Google Scholar] [CrossRef]
- Web of Science Platform. 2023. Available online: https://www-1webofscience-1com-1lbxc51ax2c09.han.amu.edu.pl/wos/woscc/analyze-results/10a4a97c-e438-47e5-8d4f-5cd4b82052df-86701a09 (accessed on 1 March 2023).
- Statistical Yearbook of the Republic of Poland. Statistics Poland. 2022. Available online: https://stat.gov.pl/ (accessed on 1 March 2023).
- Geography of Poland. Available online: https://en.wikipedia.org/wiki/Geography_of_Poland (accessed on 1 March 2023).
- File:Polska Relief.jpg. (27 September 2020). Wikimedia Commons, the Free Media Repository. Retrieved. 1 March 2023. Available online: https://commons.wikimedia.org/w/index.php?title=File:Polska_relief.jpg&oldid=473655244 (accessed on 1 March 2023).
- Climate-Data-org. Available online: https://pl.climate-data.org/europa/polska/masovian-voivodeship/warszawa-4560/ (accessed on 1 March 2023).
- Kornaś, J. Wpływ człowieka i jego gospodarki na szatę roślinną Polski. Flora synantropijna. In Szata roślinna Polski; Szafer, W., Zarzycki, K., Eds.; PWN: Warszawa, Poland, 1977; Volume 1, pp. 95–128. (In Polish) [Google Scholar]
- Symonides, E. Ochrona Przyrody; Wydawnictwo Uniwersytetu Warszawskiego: Warszawa, Poland, 2008; 766p. (In Polish) [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Vascular Plants of Poland. An Annotated Checklist; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2020; 526p. [Google Scholar]
- Zając, A.; Zając, M. (Eds.) Distribution Atlas of Vascular Plants in Poland; Laboratory of Computer Chorology, Institute of Botany, Jagiellonian University: Kraków, Poland, 2001; 716p. [Google Scholar]
- Zając, A.; Zając, M. (Eds.) Distribution Atlas of Vascular Plants in Poland. Appendix; Institute of Botany, Jagiellonian University: Kraków, Poland, 2019; 321p. [Google Scholar]
- Zarzycki, K.; Trzcińska-Tacik, H.; Różański, W.; Szeląg, Z.; Wołek, J.; Korzeniak, U. Ecological indicator values of vascular plants of Poland. In Biodiversity of Poland; Mirek, Z., Ed.; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002; Volume 1, 183p. [Google Scholar]
- Matuszkiewicz, W. Przewodnik do oznaczania zbiorowisk roślinnych Polski. In Vademecum Geobotanicum; Faliński, J., Ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2001; 537p. (In Polish) [Google Scholar]
- Kaźmierczakowa, R.; Bloch-Orłowska, J.; Celka, Z.; Cwener, A.; Dajdok, Z.; Michalska-Hejduk, D.; Pawlikowski, P.; Szczęśniak, E.; Ziarnek, K. Polska Czerwona Lista Paprotników i Roślin Kwiatowych; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2016; 44p. [Google Scholar]
- Zając, A. Pochodzenie Archeofitów Występujących w Polsce; Rozpr. Habil. Uniw. Jagiell: Kraków, Poland, 1979; Volume 29, 213p. [Google Scholar]
- Zając, M.; Zając, A.; Tokarska-Guzik, B. Extinct and endangered archaeophytes and the dynamics of their diversity in Poland. Biodivers. Res. Conserv. 2009, 13, 17–24. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B. The Establishment and Spread of Alien Plant Species (Kenophytes) in the Flora of Poland; Prace Naukowe Uniw. Śląskiego w Katowicach: Katowice, Poland, 2005; Volume 2372, pp. 1–192. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; 197p. (In Polish) [Google Scholar]
- Urbisz, A. Occurrence of Temporarily-Introduced Alien Plant Species (Ephemerophytes) in Poland—Scale and Assessment of the Phenomenon; Wyd. Uniw. Śląskiego: Katowice, Poland, 2011; 199p. [Google Scholar]
- Kaźmierczakowa, R.; Zarzycki, K.; Mirek, Z. (Eds.) Polska Czerwona Księga Roślin. Paprotniki i Rośliny Kwiatowe, 3rd ed.; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2014; 895p. (In Polish) [Google Scholar]
- Holub, J.; Jirásek, V. Zur Vereinheitlichung der terminologie in der phytogeographie. Folia Geobot. Phytotaxon. 1967, 2, 69–113. [Google Scholar] [CrossRef]
- Kornaś, J. Geograficzno-historyczna klasyfikacja roślin synantropijnych. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1968, 25, 33–41. (In Polish) [Google Scholar]
- Kornaś, J. Oddziaływanie człowieka na florę: Mechanizmy i konsekwencje. Wiadomości Bot. 1981, 25, 165–182. (In Polish) [Google Scholar]
- Schroeder, F.G. Zur Klassifizierung der Antropochoren. Vegetatio 1969, 16, 225–238. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: Towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Trzcińska-Tacik, H. Flora Synantropijna Krakowa; Rozprawy habilitacyjne UJ: Kraków, Poland, 1979; Volume 22, 278p. (In Polish) [Google Scholar]
- Mirek, Z. Problemy klasyfikacji roślin synantropijnych. Wiadomości Bot. 1981, 25, 45–54. [Google Scholar]
- Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0; IUCN: Gland, Switzerland; Cambridge, UK, 2012; 41p.
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde; Springer: New York, NY, USA, 1964; 865p. [Google Scholar]
- Jackowiak, B. Antropogeniczne przemiany flory roślin naczyniowych Poznania; Wyd. UAM, Seria Biologia: Poznań, Poland, 1990; Volume 42, 232p. (In Polish) [Google Scholar]
- Jackowiak, B. Methodological proposals for studies on the structure and dynamics of urban flora. Pol. Bot. Stud. 2006, 22, 251–260. [Google Scholar]
- Sudnik-Wójcikowska, B. Studies on Indices of Flora Synanthropization. Flora 1992, 187, 37–50. [Google Scholar] [CrossRef]
- Fukarek, F. Der Mensch beeinflusst die Pflanzenwelt. In Pflanzenwelt der Erde; Fukarek, F., Ed.; Urania Verlag: Jena, Germany; Berlin, Germany; Leipzig, Germany, 1979; pp. 65–77. [Google Scholar]
- Kornaś, J. Wymieranie flory europejskiej—Fakty, interpretacje, prognozy. Phytocoenosis 1976, 5, 173–185. (In Polish) [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen; 5. Stark Veränd. u. Verb. Aufl.; Ulmer: Stuttgart, Germany, 1996; 989p. [Google Scholar]
- Faliński, J.B. Antropogeniczna Roślinność Puszczy Białowieskiej Jako Wynik Synantropizacji Naturalnego Kompleksu Leśnego; Rozpr. Uniw. Warsz.: Warszawa, Poland, 1966; Volume 13, 256p. (In Polish) [Google Scholar]
- Faliński, J.B. Flora i roślinność synantropijna wsi i miast. Próba analizy porównawczej. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1971, 27, 15–38. [Google Scholar]
- Sukopp, H. Apophytes in the flora of Central Europe. Pol. Bot. Stud. 2006, 22, 473–485. Available online: https://www.cabdirect.org/cabdirect/abstract/20083013267 (accessed on 1 March 2023).
- Sukopp, H. Apophyten in der Flora von Mitteleuropa. Braunschw. Geobot. Arb. 2008, 9, 443–458. [Google Scholar]
- Kopecký, K. Der Apophytisierungsprozess und die Apophytengesellschaften der Galio-Urticetea mit einigen Beispielen aus der südwestlichen Umgebung von Praha. Fol. Geobot. Phytotax. 1984, 19, 113–138. [Google Scholar] [CrossRef]
- Kopecký, K. Der Apophytisierungsprozess am Beispiel der Saumgesellschaften mit Chaerophyllum aromaticum L. in der Tschechoslowakei. Tuexenia 1985, 5, 127–130. [Google Scholar]
- Kopecký, K. Einfluss der Strassen auf die Synanthropisierung der Flora und Vegetation nach Beobachtungen in der Tschechoslovakei. Folia Geobot. Phytotaxon. 1988, 23, 145–172. [Google Scholar] [CrossRef]
- Wittig, R. The origin and development of the urban flora of Central Europe. Urban Ecosyst. 2004, 7, 323–339. [Google Scholar] [CrossRef]
- Jackowiak, B. Modele Ekspansji Roślin Synantropijnych i Transgenicznych; Wydawn. Uniw. Warszawskiego: Warsaw, Poland, 1999; Volume 6, pp. 3–16. (In Polish) [Google Scholar]
- Trzcińska-Tacik, H.; Wasylikowa, K. History of the synanthropic changes of flora and vegetation of Poland. Memorab. Zool. 1982, 3, 47–69. [Google Scholar]
- Latałowa, M.; Jarosińska, J.; Badura, M. Elbląg średniowieczny w świetle dotychczasowych materiałów archeobotanicznych. Archeol. Pol. 1998, 43, 147–166. (In Polish) [Google Scholar]
- Woch, M.W. Antropofity znalezione w trakcie badań archeobotanicznych średniowiecznego Krakowa. In Rośliny w Życiu Codziennym Mieszkańców Średniowiecznego Krakowa; Mueller-Bieniek, A., Ed.; Instytut Botaniki im. W. Szafera PAN: Kraków, Poland, 2012; pp. 185–210. (In Polish) [Google Scholar]
- Tokarska-Guzik, B. Invasive ability of kenophytes occurring in Poland: A tentative assessment. In Biological Invasions—From Ecology to Control; Nentwig, N., Bacher, S., Cock, M., Dietz, H.J., Gigon, A., Wittenberg, R., Eds.; Neobiota; Pensoft Publishers: Sofia, Bulgaria, 2005; Volume 6, pp. 47–65. [Google Scholar]
- Faliński, J.B. Stadia neofityzmu i stosunek neofitów do innych komponentów zbiorowiska. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1968, 25, 15–31. (In Polish) [Google Scholar]
- Faliński, J.B. Zbiorowiska autogeniczne i antropogeniczne. Próba określenia i klasyfikacji. Dyskusje fitosocjologiczne. Ekol. Pol. Ser. B 1969, 15, 173–182. (In Polish) [Google Scholar]
- Faliński, J.B. Invasive alien plants and vegetation dynamics. In Plant Invasions: Ecological Mechanisms and Human Responses; Starfinger, U., Edwards, K., Kowarik, I., Williamson, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 3–21. [Google Scholar]
- Udvardy, M.D.F. Dynamic Zoogeography with Special Reference to Land Animals; Litton Educational Publishing, Inc.: New York, NY, USA; Van Nostrand Reinhold Company: New York, NY, USA, 1969; 445p. [Google Scholar]
- Danihelka, J.; Chrtek Jr, J.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. Available online: https://www.cabdirect.org/cabdirect/abstract/20123395489 (accessed on 1 March 2023).
- Kühn, I.; Klotz, S. The alien flora of Germany—Basics from a new German database. In Plant Invasions: Ecological Threats and Management Solutions; Child, L.E., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 90–100. [Google Scholar]
- Medvecká, J.; Kliment, J.; Májeková, J.; Halada, L.; Zaliberová, M.; Gojdičová, E.; Feráková, V.; Jarolimek, L. Inventory of the alien flora of Slovakia. Preslia 2012, 84, 257–309. Available online: https://www.cabdirect.org/cabdirect/abstract/20123299140 (accessed on 1 March 2023).
- Piotrowska, H.; Żukowski, W.; Jackowiak, B. Rośliny Naczyniowe Słowińskiego Parku Narodowego; Prace Zakładu Taksonomii Roślin UAM w Poznaniu, Bogucki Wyd. Nauk.: Poznań, Poland, 1997; Volume 6, 216p. (In Polish) [Google Scholar]
- Żukowski, W.; Latowski, K.; Jackowiak, B.; Chmiel, J. Rośliny naczyniowe Wielkopolskiego Parku Narodowego; Prace Zakładu Taksonomii Roślin UAM w Poznaniu, Bogucki Wyd. Nauk.: Poznań, Poland, 1995; Volume 4, 231p. (In Polish) [Google Scholar]
- Sudnik-Wójcikowska, B. Flora Miasta Warszawy i jej Przemiany w Ciągu XIX i XX Wieku; Wyd. UW: Warszawa, Poland, 1987; Volume 1, 242p, Volume 2, 435p. (In Polish) [Google Scholar]
- Chmiel, J. Flora Roślin Naczyniowych Wschodniej Części Pojezierza Gnieźnieńskiego i jej Antropogeniczne Przemiany; Prace Zakładu Taksonomii Roślin UAM w Poznaniu, Wyd. Sorus: Poznań, Poland, 1993; 202p. (In Polish) [Google Scholar]
- Urbisz, A. Flora naczyniowa Płaskowyżu Rybnickiego na tle antropogenicznych przemian tego obszaru. Scr. Rudensia 1996, 6, 174. (In Polish) [Google Scholar]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Delipetrou, P.; Essl, F.; Hejda, M.; Jarošík, V.; Pergl, J.; Winter, M.; Andriopoulos, P.; et al. Alien flora of Europe: Species diversity, geographical pattern and state of the art of research. Preslia 2008, 80, 101–149. Available online: http://www.preslia.cz/P082Lam.pdf (accessed on 1 March 2023).
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.C. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Protopopova, V.V.; Shevera, M.; Fedoronchuk, M.M. The geographical analysis of apophytes fraction of urban floras of Ukraine. Thaiszhia J. Bot. Kosice 2012, 22, 181–189. Available online: https://www.upjs.sk/public/media/7803/181-189-protopopova-et-al-upr.pdf (accessed on 1 March 2023).
- Zając, A. Studies on the origin of archaeophytes in Poland. Part I. Methodical consideration. Zesz. Nauk. Uniw. Jagiellońskiego Prace Bot. 1983, 11, 87–107. [Google Scholar]
- Zając, A. Studies on the origin of archaeophytes in Poland. Part II. Taxa of Mediterranean and Atlantic-Mediterranean origin. Zesz. Nauk. Uniw. Jagiellońskiego Prace Bot. 1987, 14, 7–50. [Google Scholar]
- Zając, A. Studies on the origin of archaeophytes in Poland. Part III. Taxa of Irano-Turanian, Euro-Siberian-Irano-Turanian and Mediterranean-Irano-Turanian origin. Zesz. Nauk. Uniw. Jagiellońskiego Pr. Bot. 1987, 15, 93–129. [Google Scholar]
- Zając, A. Studies on the origin of archaeophytes in Poland. Part IV. Taxa of Pontic-Pannonian, Mediterraneo-South Asiatic, South Asiatic and Middle European origin, archaeophyta anthropogena, Archaeophyta resistentia, Archaeophytes of unknown origin. Zesz. Nauk. Uniw. Jagiellońskiego Pr. Bot. 1988, 11, 87–107. [Google Scholar]
- Kornaś, J. Prowizoryczna liczba nowszych przybyszów synantropijnych (kenofitów) zadomowionych w Polsce. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1968, 25, 43–53. (In Polish) [Google Scholar]
- Eliáš, P.; Dítě, D.; Kliment, J.; Hrivnák, R.; Feráková, V. Red list of ferns and flowering plants of Slovakia, 5th edition (October 2014). Biologia 2015, 70, 218–228. [Google Scholar] [CrossRef]
- Grulich, V. Red List of vascular plants of the Czech Republic. Preslia 2012, 84, 631–645. Available online: https://www.cabdirect.org/cabdirect/abstract/20123395488 (accessed on 1 March 2023).
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M.; et al. Red list of threatened vascular plants in Italy. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- Moreno, J.C. Lista Roja 2008 de la Flora Vascular Española. Dirección General de Medio Natural y Política Forestal; Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la Conservación de Plantas: Madrid, Spain, 2008; 86p. [Google Scholar]
- Stroh, P.; Leach, S.J.; August, T.A.; Walker, K.J.; Pearman, D.A.; Rumsey, F.J.; Harrower, C.A.; Fay, M.F.; Martin, J.P.; Pankhurst, T. A vascular plant red list for England. BSBI News 2014, 9, 1–193. [Google Scholar]
- Jäger, E.J. Veränderungen des Artenbestandes von Floren unter dem Einfluß des Menschen. Biol. Rundsch. 1977, 15, 287–300. [Google Scholar]
- Jäger, E.J. Möglichkeiten der Prognose synanthroper Pflanzenausbreitungen. Flora 1988, 180, 101–131. Available online: https://www.sciencedirect.com/science/article/abs/pii/S036725301730302X (accessed on 1 March 2023). [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011; 130p. [Google Scholar]
- Le Roux, J.J.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent Anthropogenic Plant Extinctions Differ in Biodiversity Hotspots and Coldspots. Curr. Biol. 2019, 29, 2912–2918. Available online: https://www.sciencedirect.com/science/article/pii/S0960982219309431 (accessed on 1 March 2023). [CrossRef]
Figure 1.
The dynamics of published papers in the Web of Science database-indexed journals from 1980–2022 on plant extinction and invasion [43].
Figure 1.
The dynamics of published papers in the Web of Science database-indexed journals from 1980–2022 on plant extinction and invasion [43].

Figure 2.
Physical map of Poland [46].
Figure 2.
Physical map of Poland [46].

Figure 3.
Climate diagram of Warsaw [47].
Figure 3.
Climate diagram of Warsaw [47].

Figure 4.
The number of species from basic geographic-historical groups in contemporary Polish flora (N = 3614).
Figure 4.
The number of species from basic geographic-historical groups in contemporary Polish flora (N = 3614).

Figure 5.
Frequency of native and established alien species in Poland. Note: nsNS—non-synanthropic native species; frequency classes according to Zarzycki et al. [53], supplemented by the author; Fr1—very small number of stations (up to several dozen), Fr2—small number of stations (up to 100), Fr3—large number of stations, but mainly in one region, Fr4—large number of stations in many regions, Fr5—common throughout Poland.
Figure 5.
Frequency of native and established alien species in Poland. Note: nsNS—non-synanthropic native species; frequency classes according to Zarzycki et al. [53], supplemented by the author; Fr1—very small number of stations (up to several dozen), Fr2—small number of stations (up to 100), Fr3—large number of stations, but mainly in one region, Fr4—large number of stations in many regions, Fr5—common throughout Poland.

Figure 6.
The share of extinct and threatened species in the flora of Poland (N = 2141).

Figure 7.
The number of native species and archaeophytes in the different threat categories.

Figure 8.
Synecological spectrum of the Polish flora.

Figure 9.
Sequence of synecological groups ordered by the increasing proportion of apophytes, neophytes, and archaeophytes (synanthropization index of flora, SynI): nsNS—non synanthropic native species, Ap—apophytes, Ar—archaeophytes, Ne—neophytes.
Figure 9.
Sequence of synecological groups ordered by the increasing proportion of apophytes, neophytes, and archaeophytes (synanthropization index of flora, SynI): nsNS—non synanthropic native species, Ap—apophytes, Ar—archaeophytes, Ne—neophytes.

Figure 10.
Sequence of synecological groups ordered by the decreasing extinct index (ExI (%)): NotT—Not Threatened, EX—Extinct, CR—Critically Endangered, E—Endangered, V—vulnerable, NT—Near Threatened, DD—Data Deficient.
Figure 10.
Sequence of synecological groups ordered by the decreasing extinct index (ExI (%)): NotT—Not Threatened, EX—Extinct, CR—Critically Endangered, E—Endangered, V—vulnerable, NT—Near Threatened, DD—Data Deficient.

Figure 11.
Percentages of natural and anthropogenic threat factors for species in the flora of Poland by area of human activity (N = 768): natural determinants (ND), plant collecting and hoarding (CH), forest management (FM), agriculture (AG), water management (WM), settlement and urbanization (SU), investments (IN), mining (MI), tourism, recreation, and sport (TR), supra-regional environmental changes (EC).
Figure 11.
Percentages of natural and anthropogenic threat factors for species in the flora of Poland by area of human activity (N = 768): natural determinants (ND), plant collecting and hoarding (CH), forest management (FM), agriculture (AG), water management (WM), settlement and urbanization (SU), investments (IN), mining (MI), tourism, recreation, and sport (TR), supra-regional environmental changes (EC).

Figure 12.
Natural and anthropogenic factors threatening species representing the distinguished synecological groups by human activity areas. Abbreviations of the natural and anthropogenic threat factors are given in Figure 11.
Figure 12.
Natural and anthropogenic factors threatening species representing the distinguished synecological groups by human activity areas. Abbreviations of the natural and anthropogenic threat factors are given in Figure 11.


{kind=link}
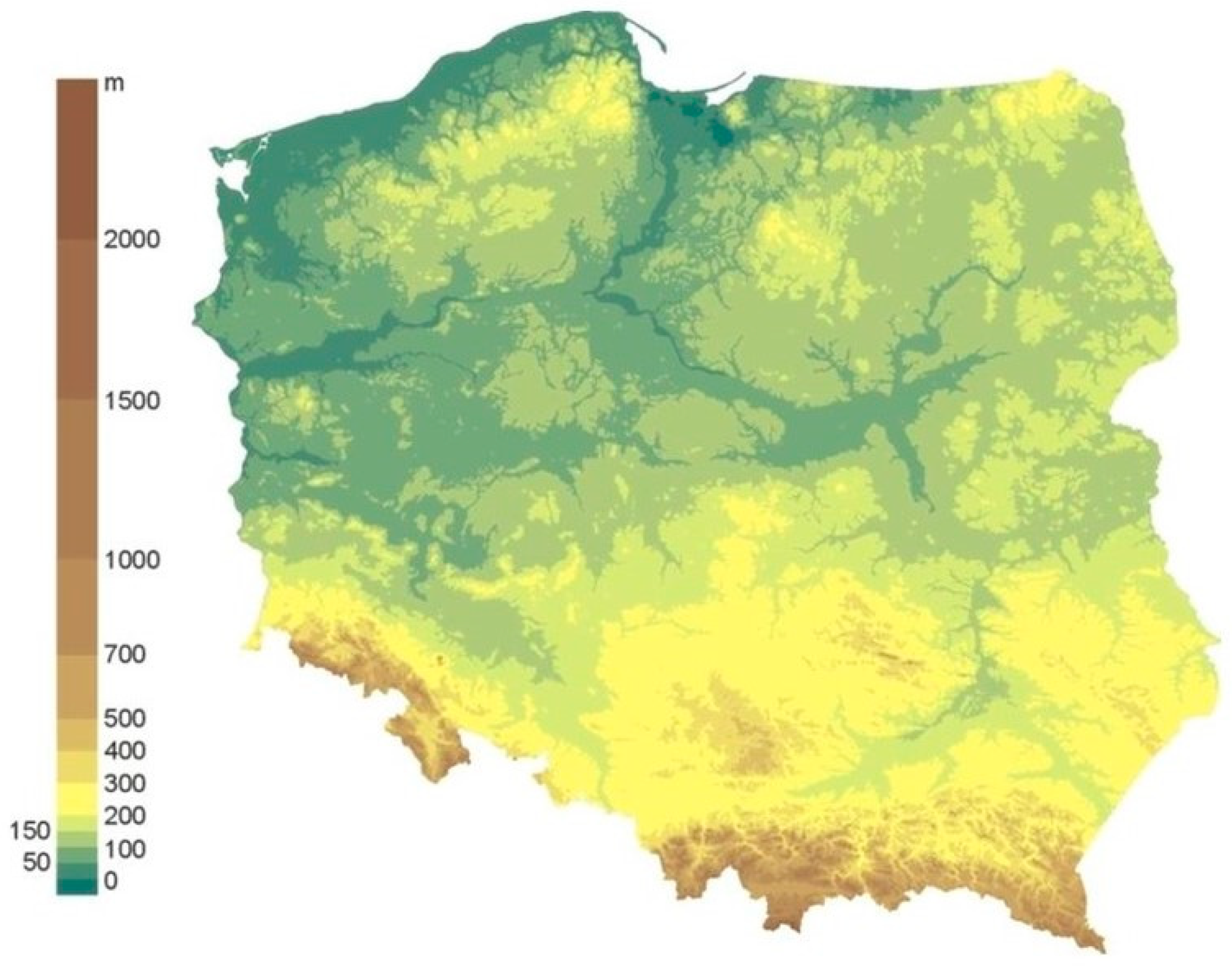{kind=link}
{kind=link}
{kind=link}
{kind=link}
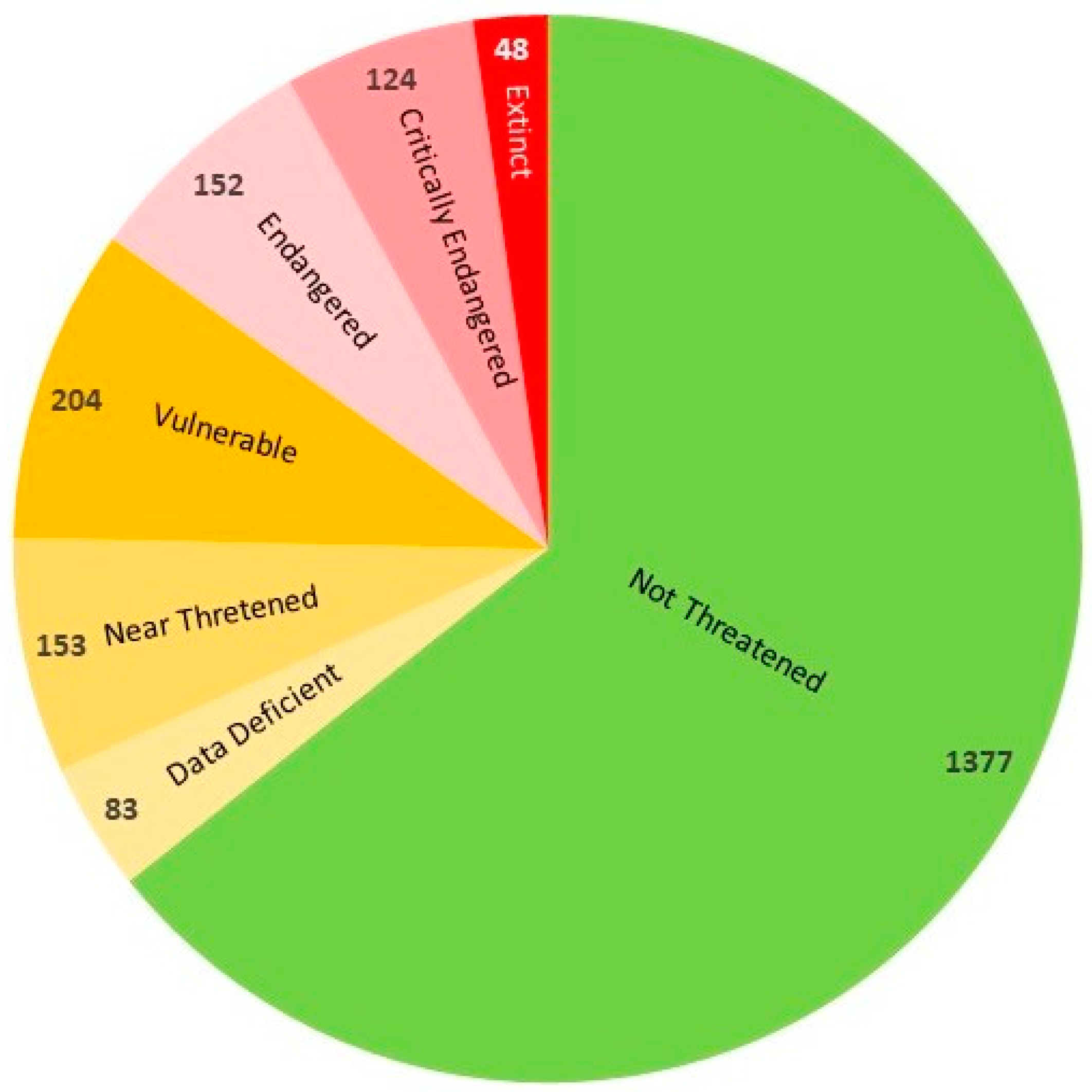{kind=link}
{kind=link}
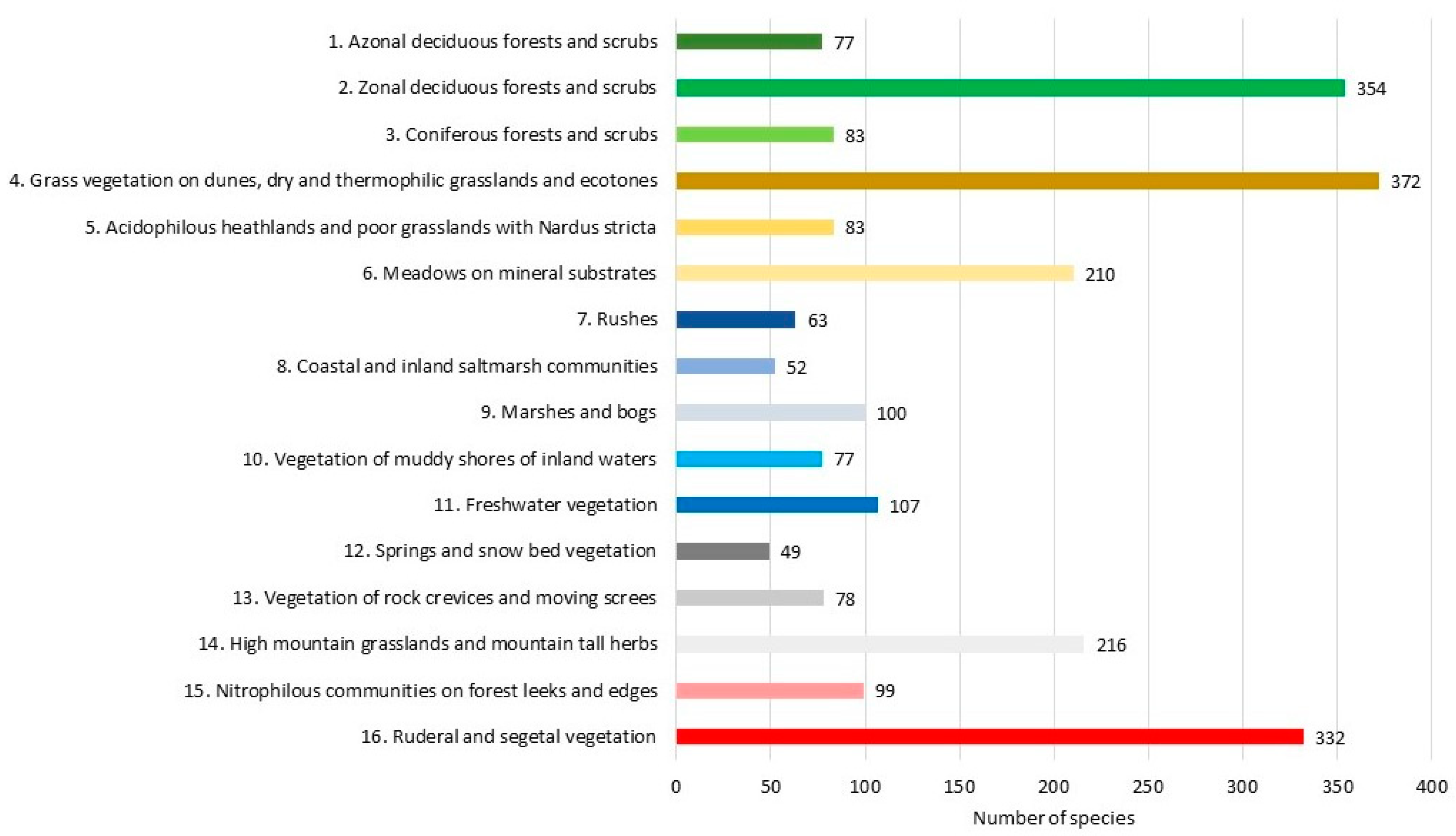{kind=link}
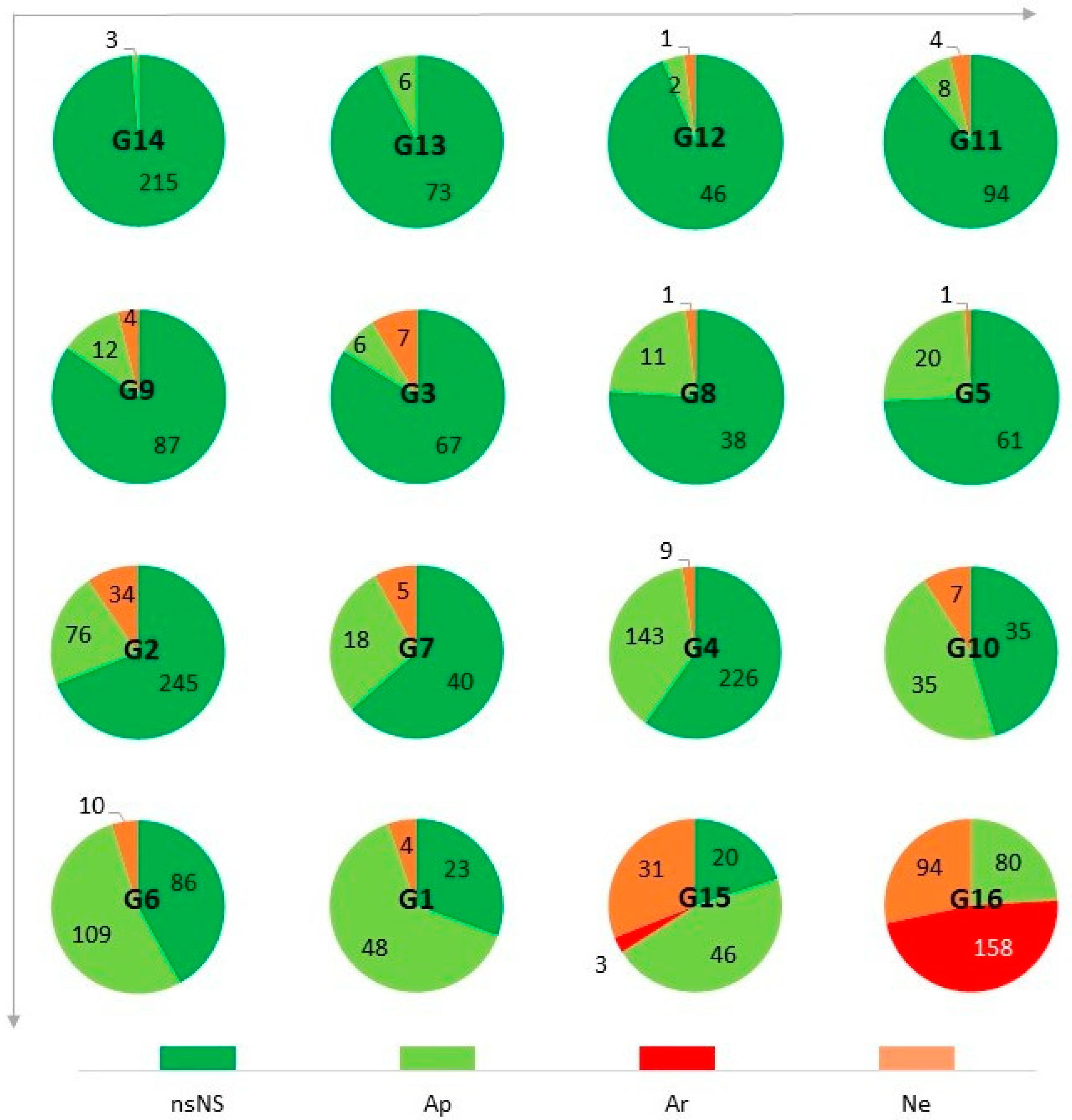{kind=link}
{kind=link}
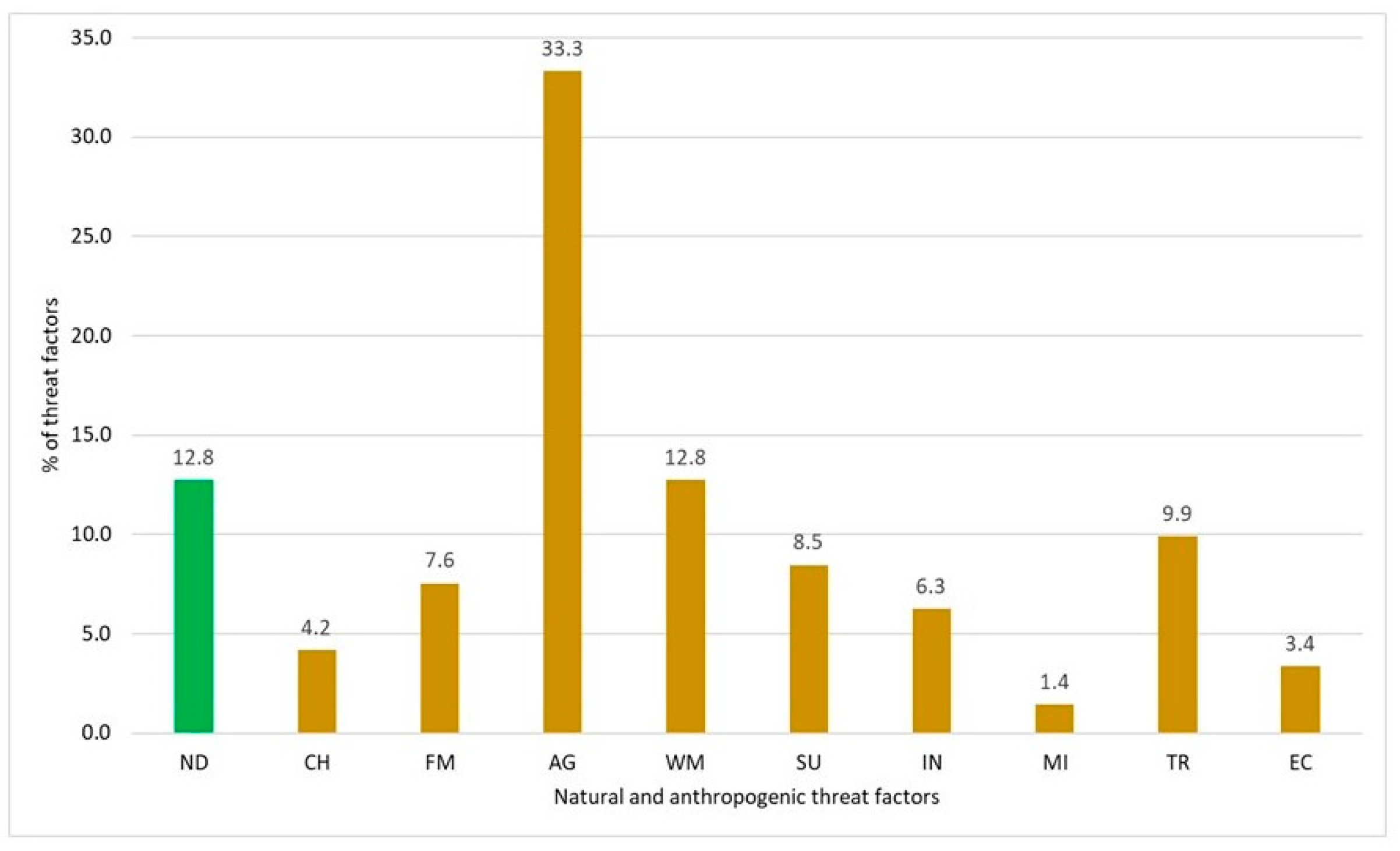{kind=link}
{kind=link}
Table 1.
The geographic-historical classification of the flora taking into account the response of species to human impacts.
Table 1.
The geographic-historical classification of the flora taking into account the response of species to human impacts.
| Category Name | Classification Criteria | Explanation |
|---|---|---|
| 1 Native plants | Taxa area of origin | Taxa that have originated in a given area without human involvement or that have arrived there without any intentional or unintentional intervention of humans from an area in which they are native [66]. |
| 1.1 Native non-synanthropic plants | Habitats occupied in the native area | Taxa whose occurrence is limited to natural and seminatural habitats. |
| 1.2 Native synanthropic plants (Apophytes) | Taxa found exclusively in man-made habitats (especially ruderal and segetal habitats) or occupying them permanently alongside natural and seminatural habitats. | |
| 2 Alien plants (Anthropophytes) | Taxa area of origin | Taxa in a given area (see below) whose presence there is due to intentional or unintentional human involvement, or which have arrived there without the help of people from an area in which they are alien [66]. |
| 2.1 Naturalized (established) plants (Metaphytes of the Trzcińska-Tacik [67] and Kornaś [64]) sense | Persistence of establishment | Permanently established anthropophytes [64,67]. Alien taxa that sustain self-replacing populations for at least 10 years without direct intervention by people (or in spite of human intervention) by recruitment from the seed or ramets (tillers, tubers, bulbs, fragments, etc.) capable of independent growth [66]. |
| 2.1.1 Archaeophytes | Time of arrival (introduction) in the study area | Alien taxa introduced more or less before the year 1500, both intentionally and accidentally, regardless of the degree of establishment (naturalization). |
| 2.1.2 Neophytes (Kenophytes of the Kornaś [64]) sense | Alien taxa introduced more or less after the year 1500, both intentionally and accidentally, regardless of the degree of establishment (naturalization). | |
| 2.1.2.1 Epecophytes | Degree of establishment (naturalization) | Neophytes established in human-made habitats. |
| 2.1.2.2 Hemiagriofity | Neophytes established in seminatural habitats. | |
| 2.1.2.3 Holoagriophytes | Neophytes established in natural habitats. | |
| 2.2 Casual alien plants (Diaphytes of the Trzcińska-Tacik [67] and Kornaś [64]) sense | Persistence of establishment | Alien plants that may flourish and even reproduce occasionally outside of cultivation in an area, but that eventually die out, because they do not form self-replacing populations and rely on repeated introductions for their persistence [66]. |
| 2.2.1 Ephemerophytes | Source and way of importation | Plants introduced into an area without intentional human involvement. |
| 2.2.2 Ergasiophytes | Alien plants escaping from crops grown in the area. |
Table 2.
Classification of plant communities.
| Number and Name of the Synecological Group |
|---|
| Description of the Synecological Group Class Names |
| 1. Azonal deciduous forests and scrubs |
| Willow-poplar forests and thickets, found in river valleys on sandy, gravelly, or stony alluviums, with poplar (Populus alba L., P. nigra L.) and willow (Salix alba L.) and deciduous forests with the prevailing alder (Alnus glutinosa (L.) Gaertn.) developing on wet peaty and peaty-mineral soils. Salicetea purpureae Moor 1958; Alnetea glutinosae Br.-Bl. et R. Tx. 1943. |
| 2. Zonal deciduous forest and scrubs Deciduous forests and scrubs shedding their leaves for the winter, found on mineral soils of varying nutrient content, moisture content and reaction. The forest stand is made up of elm (Ulmus laevis Pall., U. minor Mill., U. glabra Huds.), ash (Fraxinus excelsior L.), oak (Quercus robur L., Q. petraea (Matt.) Liebl.), lime (Tilia cordata Mill., T. platyphyllos Scop.), and beech (Fagus sylvatica L.). Querco-Fagetea Br.-Bl. et Vlieg. 1937; Quercetea robori-petraeae Br.-Bl. et R. Tx. 1943; Rhamno-Prunetea Moor 1958. |
| 3. Coniferous forests and scrubs |
| Acidophilous, oligo-, and mesotrophic communities with a predominance of coniferous species (Pinus L., Picea A. Dietr., Abies (L.) H. Karst), shrubs and abundant mosses; xerothermic forest and scrub communities dominated by pine (Pinus sylvestris L.). Vaccinio-Piceetea Br.-Bl. 1939; Erico-Pinetea Horvat 1959. |
| 4. Grass vegetation on dunes, dry and thermophilic grasslands and ecotones |
| The group encompasses a wide spectrum of vegetation; from communities of grand grasses on the coastal dunes, through low grasslands overgrowing soils with an elevated heavy metal content, dry grasslands on a mineral substrate to thermophilic grasslands, and communities forming an ecotone at the forest boundary. Ammophiletea Br.-Bl. et R. Tx. 1943; Violetea calaminariae Br.-Bl. et R.Tx. 1943; Koelerio-Corynephoretea canescentis Klika in Klika et Novak 1941; Festuco-Brometea Br.-Bl. et R.Tx. 1943; Trifolio-Geranietea sanguinei Th. Müller 1962. |
| 5. Acidophilous heathlands and poor grasslands with Nardus stricta |
| Semi-natural and anthropogenic communities, which in the primary landscape occupied small areas under specific environmental conditions and as a result of human activity have spread significantly. These include shrub communities with Calluna vulgaris (L.) Hull as well as acidophilous low grasslands and meadows with the dominant grass Nardus stricta L. Nardo-Callunetea Prsg 1949. |
| 6. Meadows on mineral substrates |
| Semi-natural and anthropogenic turf meadow and pasture communities on meso- and eutrophic undrained mineral soils. Meadows of this type also occupy organic-mineral soils and desiccated low peats. Molinio-Arrhenatheretea R.Tx. 1937. |
| 7. Rushes |
| Communities of high grasses and sedges with dicotyledonous herbaceous perennials, found in the littoral and riparian zone of the inland reservoirs of standing and flowing water. Phragmitetea R. Tx. et Prsg 1942. |
| 8. Coastal and inland saltmarsh communities |
| Extremely poor “submarine meadows” found in the littoral zone of the Baltic Sea; therophyte communities developing along the Baltic coastline; communities of the initial stages of the coastal dune formation, communities of muddy, highly saline habitats (both coastal and inland); rush-meadow communities found in the supralittoral of salt water bodies. Zosteretea marinae Pign. 1953; Rupietea maritimae J. Tx. 1960; Cakiletea maritimae R. Tx. et Prsg 1950; Thero-Salicornietea Pign. 1953 em. R. Tx. 1954 in R. Tx. et Oberd. 1958; Asteretea tripolium Westh. et Beeft. ap. Beeft 1962; Ammophiletea Br.-Bl. et R. Tx. 1943 (Agropyro-Honckenyion R. Tx. 1952). |
| 9. Marshes and bogs |
| Wet heath and raised bogs on acidic and dystrophic habitats, fed exclusively or predominantly by rainwater, as well as low-sedge swamp meadow communities, transitional and low bogs. Oxycocco-Sphagnetea Br.-Bl. et R. Tx. 1943; Scheuchzerio-Caricetea nigrae (Nordh. 1937) R. Tx. 1937. |
| 10. Vegetation of muddy shores of the inland waters |
| Moderately nitrophilous, mostly natural summer and autumn therophyte communities found on the drying shores of the inland water bodies. Bidentetea tripartiti R. Tx., Lohm. et Prsg. 1950; Isoëto-Nanojuncetea Br.-Bl. et R. Tx. 1943. |
| 11. Freshwater vegetation |
| Communities of water reefs (Lemna L.) forming aggregations on the surface of still and slow-moving waters, as well as communities of small perennials and macrophytes found in the waters with a wide range of nutrients. Lemnetea R. Tx. 1955; Potametea R. Tx. et Prsg; Littorelletea Br.-Bl. et R. Tx. 1943; Utricularietea intermedio-minoris Den Hartog et Segal 1964 em. Pietsch 1965. |
| 12. Springs and snow bed vegetation |
| Spring communities, often with a significant proportion of bryophytes, as well as high-mountain snow beds communities, i.e., places with a shortened growing season due to prolonged snow. Montio-Cardaminetea Br.-Bl. et R. Tx. 1943; Salicetea herbaceae Br.-Bl. et al. 1947. |
| 13. Vegetation of rock crevices and moving screes |
| Natural communities of rock crevices in the mountains and anthropogenic communities in the secondary habitats (walls, ruins), as well as pioneer communities of mobile or poorly consolidated scree in the rocky parts of mountains or in the mountainous sections of river valleys. Asplenietea rupestria Br.-Bl. 1934 in Meier et Br.-Bl. 1934; Thlaspietea rotundifolii Br.-Bl. in Br.-Bl. et Jenny 1926 em. Seibert 1977. |
| 14. High mountain grasslands and mountain tall herbs |
| High mountain grasslands on calciophilic and acidophilic substrates, as well as high mountain herb and grassland. The latter are the communities of tall dicotyledonous perennials or grasses found in relatively fertile moist or wet habitats. |
| Seslerietea variae Br.-Bl. 1948 em. Oberd. 1978; Juncetea trifidi Hadač in Klika et Hadač 1944; Betulo-Adenostyletea Br.-Bl. 1948 |
| 15. Nitrophilous communities on forest leeks and edges |
| Nitrophilous communities of therophytes, perennials, and shrubs that initiate forest regeneration after the removal or destruction of a forest stand, as well as natural and semi-natural nitrophilous communities in fertile fresh, moist or wet habitats, forming ecotones at the contact between woody and herbaceous communities. Epilobietea angustifolii R. Tx. et Prsg. 1950; Artemisietea vulgaris Lohm., Prsg et R. Tx. in R. Tx. 1950; (Galio-Urticenea Pass. 1967). |
| 16. Ruderal and segetal vegetation |
| Anthropogenic weed communities of cultivated fields and annual and perennial plants overgrowing the ruderal habitats (roadsides, balks, trackways, rubble pits, etc.), as well as thermophilic rhizomatous plant communities found on the mineral grounds. Stellarietea mediae R. Tx., Lohm. et Prsg. 1950; Agropyretea intermedio-repentis (Oberd. et al. 1967) Müller et Görs 1969; Artemisietea vulgaris Lohm., Prsg. et R. Tx. in R. Tx. 1950 (Artemisienea vulgaris Th. Müller 1981 in Oberd. 1983). |
Table 3.
Indices of anthropogenic changes in the flora of synecological groups.
| SynI (%) | ApI (%) | AnI (%) | ArI (%) | NeI (%) | ExI (%) | |
|---|---|---|---|---|---|---|
| G1 | 69.3 | 64.0 | 5.3 | 0.0 | 5.3 | 4.2 |
| G2 | 31.0 | 21.4 | 9,6 | 0.0 | 9.6 | 20.9 |
| G3 | 16.3 | 7.5 | 8.8 | 0.0 | 8.8 | 37.0 |
| G4 | 40.2 | 37.8 | 2.4 | 0.0 | 2.4 | 49.1 |
| G5 | 25.6 | 24.4 | 1.2 | 0.0 | 1.2 | 37.0 |
| G6 | 58.0 | 53.2 | 4.9 | 0.0 | 4.9 | 27.7 |
| G7 | 36.5 | 28.6 | 7.9 | 0.0 | 7.9 | 31.0 |
| G8 | 24.0 | 22.0 | 2.0 | 0.0 | 2.0 | 79.6 |
| G9 | 15.5 | 11.7 | 3.9 | 0.0 | 3.9 | 69.7 |
| G10 | 54.5 | 45.5 | 9.1 | 0.0 | 9.1 | 51.4 |
| G11 | 11.3 | 7.5 | 3.8 | 0.0 | 3.8 | 53.9 |
| G12 | 6.1 | 4.1 | 2.0 | 0.0 | 2.0 | 31.3 |
| G13 | 7.6 | 7.6 | 0.0 | 0.0 | 0.0 | 40.5 |
| G14 | 1.4 | 1.4 | 0.0 | 0.0 | 0.0 | 32.1 |
| G15 | 80.0 | 46.0 | 34.0 | 3.0 | 31.0 | 11.6 |
| G16 | 100.0 | 24.1 | 75.9 | 47.6 | 28.3 | 25.1 |
| AVG | 36.1 | 25.4 | 10.7 | 3.2 | 7.5 | 37.6 |
| STD | 29.0 | 19.1 | 19.2 | 11.9 | 9.2 | 19.7 |
Note: G1–G16—synecological groups (cf. Table 2); SynI—synanthropization index of flora, ApI—apophytization index of flora, AnI—anthropophytization index of flora, ArI—archaeophytization index of flora, NeI—neophytization index of flora, ExI—extinction index of flora (cf. Section 2.3).
Table 4.
The native species groups distinguished for their adaptability to man-made habitats.
| Example Species | Threat Category | Freq. Class | Synecol. Group | Geo-Hist. Status |
|---|---|---|---|---|
| A. Species occurring only in natural and semi-natural communities | ||||
| Osmunda regalis L. | VU | 3 | 1 | nsNS |
| Corydalis pumila (Host) Rchb. | VU | 2 | 2 | nsNS |
| Neottianthe cucullata (L.) Schltr. | CR | 1 | 3 | nsNS |
| Hypericum elegans Stephan ex Willd. | CR | 1 | 4 | nsNS |
| Spiranthes spiralis (L.) Chevall. | CR | 1 | 5 | nsNS |
| Gladiolus paluster Gaudin | CR | 1 | 6 | nsNS |
| Apium nodiflorum (L.) Lag. | CR | 1 | 7 | nsNS |
| Ruppia maritima L. | VU | 1 | 8 | nsNS |
| Schoenus nigricans L. | EN | 1 | 9 | nsNS |
| Apium repens (Jacq.) Lag. | CR | 1 | 10 | nsNS |
| Isoëtes echinospora Durieu | CR | 1 | 11 | nsNS |
| Cochlearia polonica E. Fröhl. | EW | 1 | 12 | nsNS |
| Asplenium adiantum-nigrum L. | EN | 1 | 13 | nsNS |
| Androsace obtusifolia All. | EN | 1 | 14 | nsNS |
| Bromus ramosus Huds. | VU | 2 | 15 | nsNS |
| Carex elongata L. | NotT | 4 | 1 | nsNS |
| Pulmonaria officinalis L. s. s. | NotT | 2 | 2 | nsNS |
| Luzula sylvatica (Huds.) Gaudin | NotT | 3 | 3 | nsNS |
| Eryngium campestre L. | NotT | 2 | 4 | nsNS |
| Galium saxatile L. | NotT | 2 | 5 | nsNS |
| Cirsium canum (L.) All. | NotT | 3 | 6 | nsNS |
| Carex cespitosa L. | NotT | 4 | 7 | nsNS |
| Festuca salina Natho & Stohr | NotT | 1 | 8 | nsNS |
| Carex flava L. | NotT | 4 | 9 | nsNS |
| Peplis portula L. | NotT | 4 | 10 | nsNS |
| Najas major All. | NotT | 1 | 11 | nsNS |
| Stellaria uliginosa Murray | NotT | 4 | 12 | nsNS |
| Cystopteris sudetica A. Braun & Milde | NotT | 2 | 13 | nsNS |
| Gentiana frigida Haenke | NotT | 1 | 14 | nsNS |
| Bromus benekenii (Lange) Trimen | NotT | 3 | 15 | nsNS |
| B. Species occurring in natural and semi-natural communities and sporadically found in anthropogenic habitats | ||||
| Artemisia pontica L. | CR | 1 | 4 | nsNS |
| Asplenium septentrionale (L.) Hoffm. | VU | 2 | 13 | nsNS |
| Carex buekii Wimm. | NT | 1 | 7 | nsNS |
| Carex secalina Wahlenb. | CR | 1 | 8 | nsNS |
| Cirsium decussatum Janka | VU | 1 | 6 | nsNS |
| Orchis pallens L. | VU | 2 | 2 | nsNS |
| Phelipanche arenaria (Borkh.) Pomel | CR | 1 | 4 | nsNS |
| Sagina ciliata Fr. | CR | 1 | 10 | nsNS |
| C. Species occurring in natural and semi-natural communities and permanently established in anthropogenic habitats | ||||
| Plantago coronopus L. | CR | 1 | 8 | Ap |
| Spergularia salina J. Presl & C. Presl | VU | 3 | 8 | Ap |
| Triglochin maritimum L. | VU | 2 | 8 | Ap |
| Aegopodium podagraria L. | NotT | 5 | 2 | Ap |
| Asplenium ruta-muraria L. | NotT | 3 | 13 | Ap |
| Bidens tripartita L. | NotT | 5 | 10 | Ap |
| Calamagrostis epigejos (L.) Roth | NotT | 5 | 15 | Ap |
| Campanula rapunculoides L. | NotT | 5 | 4 | Ap |
| Deschampsia caespitosa (L.) P. Beauv. | NotT | 5 | 6 | Ap |
| Epilobium hirsutum L. | NotT | 5 | 7 | Ap |
| Epilobium obscurum Schreb. | NotT | 3 | 12 | Ap |
| Juncus articulatus L. em. K. Richt. | NotT | 5 | 9 | Ap |
| Lemna minor L. | NotT | 5 | 11 | Ap |
| Luzula campestris (L.) DC. | NotT | 5 | 5 | Ap |
| Solanum dulcamara L. | NotT | 5 | 1 | Ap |
| Sorbus aucuparia L. em. Hedl. | NotT | 5 | 3 | Ap |
| Thalictrum aquilegiifolium L. | NotT | 4 | 14 | Ap |
| Verbascum densiflorum Bertol. | NotT | 4 | 16 | Ap |
| D. Species occurring exclusively in man-made habitats, lacking contact with natural and semi-natural communities | ||||
| Arabidopsis thaliana (L.) Heynh. | NotT | 4 | 16 | Ap-AF |
| Arnoseris minima (L.) Schweigg. & Körte | NotT | 4 | 16 | Ap-AF |
| Berteroa incana (L.) DC. | NotT | 4 | 16 | Ap-RC |
| Melilotus officinalis (L.) Pall | NotT | 5 | 16 | Ap-RC |
Note: Threat category: CR—Critically Endangered, EN—Endangered, VU—Vulnerable, NT—Near Threatened (NT), DD—Data Deficiency, NotT—Not Threatened; Frequency class: compare Figure 5; Synecological groups: compare Table 2; Geographic-historical status: nsNS—Non-synanthropic native species, Ap—apophytes, Ap-AF—apophytes found mainly in arable fields; Ap-RC—apophytes found in ruderal habitats.
Table 5.
The alien species groups distinguished according to persistence of establishment and degree of naturalization.
Table 5.
The alien species groups distinguished according to persistence of establishment and degree of naturalization.
| Example Species | Threat Category | Freq. Class | Synecol. Group | Geo-Hist. Status |
|---|---|---|---|---|
| A. Casual alien species | ||||
| Aegilops ligustica Asch. & Graebn. | nc | 1 | nc | Cas-Eh |
| Amaranthus deflexus L. | nc | 1 | nc | Cas-Eh |
| Bidens pilosa L. | nc | 1 | nc | Cas-Eh |
| Citrus aurantium L. | nc | 1 | nc | Cas-Eh |
| Rumex triangulivalvis (Danser) Rech.f. | nc | 1 | nc | Cas-Eh |
| Sideritis montana L. | nc | 1 | nc | Cas-Eh |
| Consolida orientalis (J. Gay) Schrödinger | nc | 1 | nc | Cas-Er |
| Corylus colurna L. | nc | 1 | nc | Cas-Er |
| Cynosurus echinatus L. | nc | 1 | nc | Cas-Er |
| Daucus aureus Desf. | nc | 1 | nc | Cas-Er |
| Glaucium flavum Crantz | nc | 1 | nc | Cas-Er |
| B. Established alien species | ||||
| B1. Archaeophytes | ||||
| Camelina alyssum (Mill.) Thell. | EX | 2 | 16 | Ar-AF |
| Caucalis platycarpos L. | CR | 2 | 16 | Ar-AF |
| Ranunculus arvensis L. | EN | 3 | 16 | Ar-AF |
| Atriplex rosea L. | VU | 2 | 16 | Ar-RC |
| Chenopodium murale L. | EN | 3 | 16 | Ar-RC |
| Chenopodium vulvaria L. | EN | 3 | 16 | Ar-RC |
| Apera spica-venti (L.) P. Beauv. | NotT | 5 | 16 | Ar-AF |
| Consolida regalis Gray | NotT | 4 | 16 | Ar-AF |
| Papaver rhoeas L. | NotT | 4 | 16 | Ar-AF |
| Lamium album L. | NotT | 5 | 16 | Ar-RC |
| Artemisia absinthium L. | NotT | 4 | 16 | Ar-RC |
| Verbena officinalis L. | NotT | 4 | 16 | Ar-RC |
| B2. Neophytes | ||||
| Cochlearia danica L. | nc | nc | nc | Ep-21 |
| Duchesnea indica (Andrews) Focke | nc | nc | nc | Ep-21 |
| Senecio inaequidens DC. | nc | nc | nc | Ep-21 |
| Anthoxanthum aristatum Boiss. | nc | 3 | 16 | Ep-AF |
| Oxalis dillenii Jacq. | nc | 2 | 16 | Ep-AF |
| Silene dichotoma Ehrh. | nc | 3 | 16 | Ep-AF |
| Vicia dasycarpa Ten. | nc | 3 | 16 | Ep-AF |
| Vicia grandiflora Scop. | nc | 4 | 16 | Ep-AF |
| Amaranthus blitoides S. Watson | nc | 2 | 16 | Ep-RC |
| Atriplex tatarica L. | nc | 2 | 16 | Ep-RC |
| Iva xanthiifolia Nutt. | nc | 3 | 16 | Ep-RC |
| Lepidium virginicum L. | nc | 3 | 16 | Ep-RC |
| Reseda luteola L. | nc | 3 | 16 | Ep-RC |
| Amaranthus chlorostachys Willd. | nc | 2 | 16 | Ep-AR |
| Amaranthus retroflexus L. | nc | 5 | 16 | Ep-AR |
| Galinsoga ciliata (Raf.) S. F. Blake | nc | 4 | 16 | Ep-AR |
| Galinsoga parviflora Cav. | nc | 5 | 16 | Ep-AR |
| Oxalis corniculata L. | nc | 3 | 16 | Ep-AR |
| Aronia x prunifolia (Marshall) Rehder | nc | 1 | 9 | He-Ag |
| Clematis vitalba L. | nc | 3 | 2 | He-Ag |
| Genistella sagittalis (L.) Gams in Hegi | nc | 2 | 5 | He-Ag |
| Lactuca tatarica (L.) C. A. Mey. | nc | 1 | 8 | He-Ag |
| Lemna turionifera Landolt | nc | 2 | 11 | He-Ag |
| Onobrychis viciifolia Scop. | nc | 3 | 4 | He-Ag |
| Quercus rubra L. | nc | 4 | 3 | He-Ag |
| Rumex longifolius DC. | nc | 1 | 7 | He-Ag |
| Solidago canadensis L. | nc | 4 | 15 | He-Ag |
| Veronica filiformis Sm. | nc | 3 | 6 | He-Ag |
| Xanthium albinum (Widder) H. Scholz | nc | 3 | 10 | He-Ag |
| Acer negundo L. | nc | 4 | 1 | Ho-Ag |
| Acorus calamus L. | nc | 5 | 7 | Ho-Ag |
| Bidens frondosa L. | nc | 3 | 10 | Ho-Ag |
| Elodea canadensis Michx. | nc | 5 | 11 | Ho-Ag |
| Impatiens parviflora DC. | nc | 4 | 2 | Ho-Ag |
| Mimulus moschatus Douglas ex Lindl. | nc | 2 | 12 | Ho-Ag |
| Padus serotina (Ehrh.) Borkh. | nc | 4 | 3 | Ho-Ag |
| Spiraea tomentosa L. | nc | 2 | 6 | Ho-Ag |
Note: nc—non-classified; Threat category: EX—Extinct, CR—Critically Endangered, EN—Endangered, VU—Vulnerable; Frequency class: compare Figure 5; Synecological groups: compare Table 2; Geographic-historical status: Cas-Eh—casual alien species introduced into an area without intentional human involvement; Cas-Er—casual alien species escaping from crops grown in the area; Ar-AF—archeophytes found mainly in arable fields; Ar-RC—archeophytes found mainly in ruderal habitats; Ep-21—neophytes (epecophytes) recently established, in the 21st century; Ep-AF—neophytes (epecophytes) found mainly in arable fields; Ep-RC—neophytes (epecophytes) found mainly in ruderal habitats; Ep-AR—neophytes (epecophytes) established in arable fields and ruderal habitats; He-Ag—neophytes (hemiagriophytes) established in seminatural communities; Ho-Ag—neophytes (holoagriophytes) established in natural communities.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jackowiak, B. Man-Made Changes in the Flora and Vegetation of Poland: Current Review. Diversity 2023, 15, 618. https://doi.org/10.3390/d15050618
AMA Style
Jackowiak B. Man-Made Changes in the Flora and Vegetation of Poland: Current Review. Diversity. 2023; 15(5):618. https://doi.org/10.3390/d15050618
Chicago/Turabian StyleJackowiak, Bogdan. 2023. "Man-Made Changes in the Flora and Vegetation of Poland: Current Review" Diversity 15, no. 5: 618. https://doi.org/10.3390/d15050618
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.