Gummy Stem Blight Resistance in Melon: Inheritance Pattern and Development of Molecular Markers

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Inheritance of GSB Resistance in Melon
2.2. Selection of GSB Resistance Genes
2.3. Cloning and Sequencing of GSB Candidate Gene
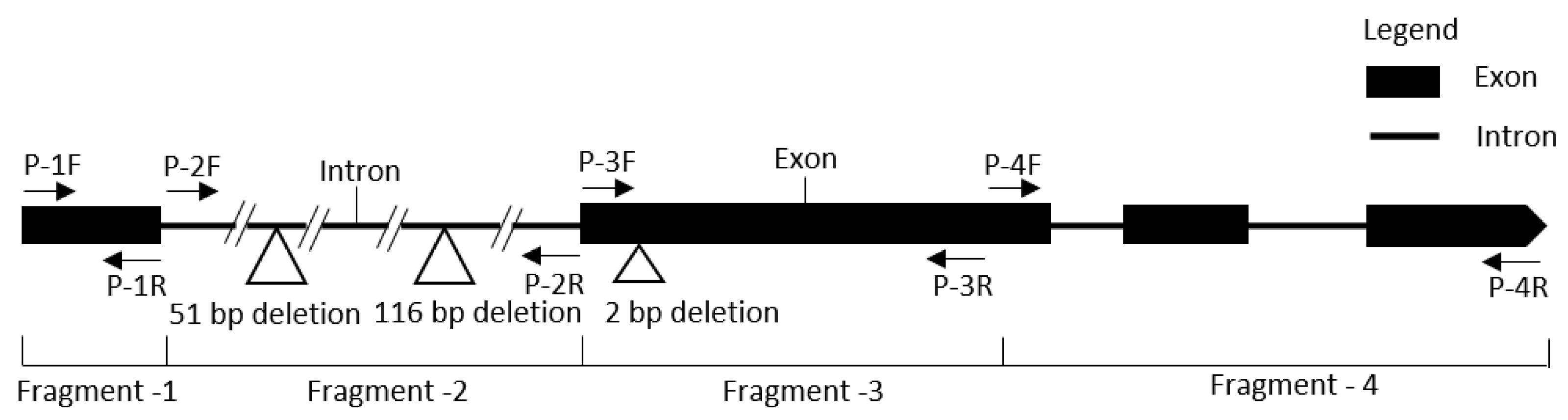
2.4. InDel Detection and Marker Development
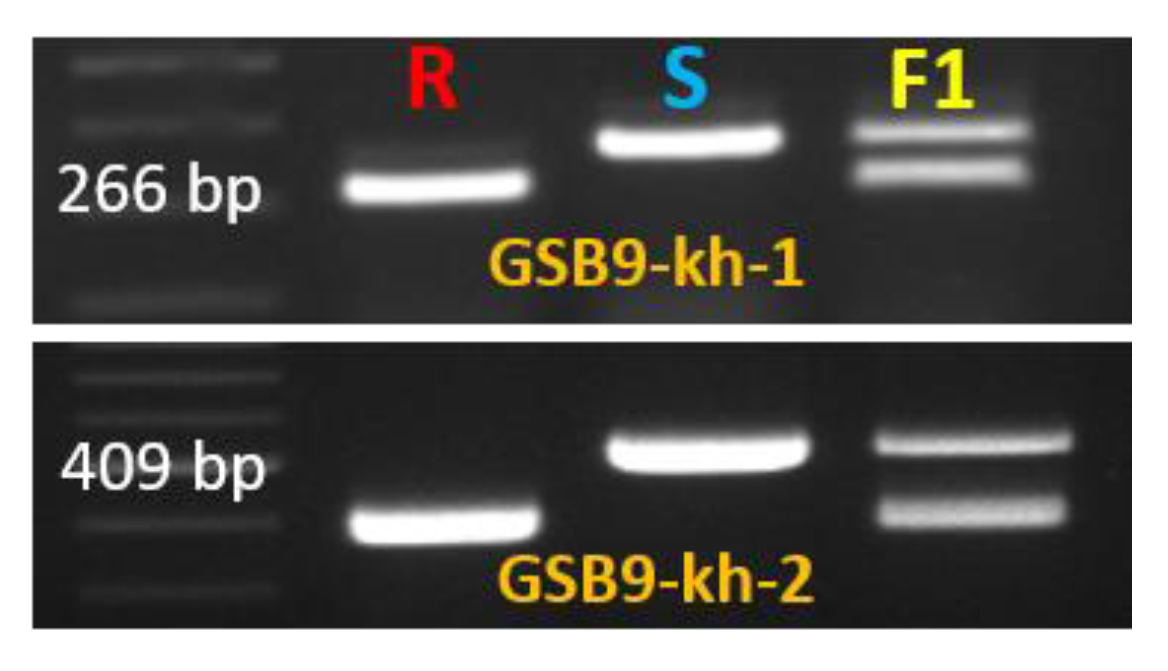
2.5. Validation of the InDel Marker
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Development of F2 Population to Explore Inheritance Patterns
4.3. Fungal Isolate and Inoculum Preparation
4.4. Inoculation Test for GSB Resistance
4.5. Disease Ratings
4.6. Exploring GSB Resistance Genes and Primer Design
4.7. DNA Extraction
4.8. PCR Conditions and Electrophoresis
4.9. Cloning and Sequencing
4.10. Statistical Analysis and Software Used
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McGrath, D.; Vawdrey, L.; Walker, I. Resistance to gummy stem blight in muskmelon. HortScience 1993, 28, 930–931. [Google Scholar]
- Lester, G.E. Antioxidant, sugar, mineral, and phytonutrient concentrations across edible fruit tissues of orange-fleshed honeydew melon (Cucumis melo L.). J. Agric. Food Chem. 2008, 56, 3694–3698. [Google Scholar] [CrossRef] [PubMed]
- Lester, G.E.; Hodges, D.M. Antioxidants associated with fruit senescence and human health: Novel orange-fleshed non-netted honey dew melon genotype comparisons following different seasonal productions and cold storage durations. Postharvest Biol. Technol. 2008, 48, 347–354. [Google Scholar] [CrossRef]
- Keinath, A.; Farnham, M.; Zitter, T. Morphological, pathological, and genetic differentiation of Didymella bryoniae and Phoma spp. isolated from cucurbits. Phytopathology 1995, 85, 364–369. [Google Scholar] [CrossRef]
- Zhang, N.; Xu, B.-H.; Bi, Y.-F.; Lou, Q.-F.; Chen, J.-F.; Qian, C.-T.; Zhang, Y.-B.; Yi, H.-P. Development of a muskmelon cultivar with improved resistance to gummy stem blight and desired agronomic traits using gene pyramiding. Czech J. Genet. Plant Breed. 2017, 53, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Sitterly, R.W.; Keinath, A.P. Gummy stem blight. In Compendium of Cucurbit Diseases; Zitter, T.A., Hopkins, D.L., Thomas, C., Eds.; APS Press: St. Paul, MN, USA, 1996; pp. 27–28. [Google Scholar]
- Wolukau, J.N.; Zhou, X.; Chen, J. Identification of amplified fragment length polymorphism markers linked to gummy stem blight (didymella bryoniae) resistance in melon (Cucumis melo L.) pi 420145. HortScience 2009, 44, 32–34. [Google Scholar]
- Norton, J.D. Gulfcoast: A Sweet Cantaloupe for the Produce Chain Store Market; Agricultural Experiment Station, Auburn University: Auburn, OR, USA, 1971. [Google Scholar]
- Zhang, Y.; Kyle, M.; Anagnostou, K.; Zitter, T.A. Screening melon (Cucumis melo) for resistance to gummy stem blight in the greenhouse and field. HortScience 1997, 32, 117–121. [Google Scholar]
- Zuniga, T.; Jantz, J.; Zitter, T.; Jahn, M. Monogenic dominant resistance to gummy stem blight in two melon (Cucumis melo) accessions. Plant Dis. 1999, 83, 1105–1107. [Google Scholar] [CrossRef]
- Sakata, Y.; Sugiyama, M.; Ohara, T.; Ishiuchi, D.; Kojima, A.; Wako, T. Identification of Melon Accessions Resistant to Gummy Stem Blight and Genetic Analysis of the Resistance Using an Efficient Technique for Seedling Test. Acta Hortic. 2002, 588, 161–164. [Google Scholar]
- Frantz, J.; Jahn, M. Five independent loci each control monogenic resistance to gummy stem blight in melon (Cucumis melo L.). Theor. Appl. Genet. 2004, 108, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Deng, G.; Mou, H.; Xu, Y.; Chen, L.; Yang, J.; Zhang, M. A re-sequencing-based ultra-dense genetic map reveals a gummy stem blight resistance-associated gene in Cucumis melo. DNA Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, Y.; Zhou, X.; CHEN, J.-F. Ssr marker linked to gummy stem blight resistance gene gsb-1 in melon and its allelism with resistance gene from PI420145. China Cucurbits Veg. 2009, 5, 1–4. [Google Scholar]
- Das, S.; Upadhyaya, H.D.; Bajaj, D.; Kujur, A.; Badoni, S.; Kumar, V.; Tripathi, S.; Gowda, C.L.; Sharma, S.; Singh, S. Deploying qtl-seq for rapid delineation of a potential candidate gene underlying major trait-associated QTL in chickpea. DNA Res. 2015, 22, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Fukino, N.; Yoshioka, Y.; Sakata, Y.; Matsumoto, S. Construction of an intervarietal genetic map of cucumber and its comparison with the melon genetic map. In Proceedings of the Cucurbitaceae 2010, Charleston, SC, USA, 14–18 November 2010; pp. 22–25. [Google Scholar]
- Li, D.; Cuevas, H.E.; Yang, L.; Li, Y.; Garcia-Mas, J.; Zalapa, J.; Staub, J.E.; Luan, F.; Reddy, U.; He, X. Syntenic relationships between cucumber (Cucumis sativus L.) and melon (C. melo L.) chromosomes as revealed by comparative genetic mapping. BMC Genom. 2011, 12, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Shi, Y.; Miao, H.; Wang, M.; Li, B.; Gu, X.; Zhang, S. Genetic analysis and QTL mapping of resistance to gummy stem blight in Cucumis sativus seedling stage. Plant Dis. 2017, 101, 1145–1152. [Google Scholar] [CrossRef]
- Brotman, Y.; Normantovich, M.; Goldenberg, Z.; Zvirin, Z.; Kovalski, I.; Stovbun, N.; Doniger, T.; Bolger, A.M.; Troadec, C.; Bendahmane, A. Dual resistance of melon to Fusarium oxysporum races 0 and 2 and to papaya ring-spot virus is controlled by a pair of head-to-head-oriented NB-LRR genes of unusual architecture. Mol. Plant 2013, 6, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Marone, D.; Russo, M.A.; Laidò, G.; De Leonardis, A.M.; Mastrangelo, A.M. Plant nucleotide binding site–leucine-rich repeat (NBS-LRR) genes: Active guardians in host defense responses. Int. J. Mol. Sci. 2013, 14, 7302–7326. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Yuan, W.; Bo, K.; Shen, J.; Pang, X.; Chen, J. Genome-wide analysis of nbs-encoding disease resistance genes in Cucumis sativus and phylogenetic study of NBS-encoding genes in cucurbitaceae crops. BMC Genom. 2013, 14, 109. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR–encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [PubMed]
- Monosi, B.; Wisser, R.; Pennill, L.; Hulbert, S. Full-genome analysis of resistance gene homologues in rice. Theor. Appl. Genet. 2004, 109, 1434–1447. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, S.; Kim, H.-T.; Thamilarasan, S.K.; Veerappan, K.; Park, J.-I.; Nou, I.-S. Whole genome re-sequencing and characterization of powdery mildew disease-associated allelic variation in melon. PLoS ONE 2016, 11, e0157524. [Google Scholar] [CrossRef] [PubMed]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- DeYoung, B.J.; Innes, R.W. Plant NBS-LRR proteins in pathogen sensing and host defense. Nat. Immunol. 2006, 7, 1243–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Wang, P.; Li, C.; Han, S.; Zhao, C.; Xia, H.; Bi, Y.; Guo, B.; Zhang, X.; Wang, X. Comparative analysis of NBS-LRR genes and their response to aspergillus flavus in arachis. PLoS ONE 2017, 12, e0171181. [Google Scholar] [CrossRef] [PubMed]
- Lorang, J.M.; Sweat, T.A.; Wolpert, T.J. Plant disease susceptibility conferred by a “resistance” gene. Proc. Natl. Acad. Sci. USA 2007, 104, 14861–14866. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Choi, W.; Thomas, C.E.; Dean, R.A. Cloning of disease-resistance homologues in end sequences of bac clones linked to fom-2, a gene conferring resistance to fusarium wilt in melon (Cucumis melo L.). Genome 2002, 45, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Hwang, J.-H.; Han, D.-Y.; Park, M.; Kim, S.; Choi, D.; Kim, Y.; Lee, G.P.; Kim, S.-T.; Park, Y.-H. Major quantitative trait loci and putative candidate genes for powdery mildew resistance and fruit-related traits revealed by an intraspecific genetic map for watermelon (Citrullus lanatus var. lanatus). PLoS ONE 2015, 10, e0145665. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Q.; Liu, Y.; Zhuang, M.; Yang, Y.; Xie, B.; Liu, B. Mapping and analysis of a novel candidate fusarium wilt resistance gene foc1 in Brassica oleracea. BMC Genom. 2014, 15, 1094. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kwon, S.I.; Saha, D.; Anyanwu, N.C.; Gassmann, W. Resistance to the pseudomonas syringae effector hopa1 is governed by the TIR-NBS-LRR protein RPS6 and is enhanced by mutations in SRFR1. Plant Physiol. 2009, 150, 1723–1732. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Shoji, H.; Ando, S.; Kanayama, Y.; Kusano, T.; Takeshita, M.; Suzuki, M.; Masuta, C. Rcy1-mediated resistance to cucumber mosaic virus is regulated by lrr domain-mediated interaction with CMV (Y) following degradation of RCY1. Mol. Plant-Microbe Interact. 2012, 25, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Afrin, K.S.; Rahim, M.A.; Park, J.-I.; Natarajan, S.; Kim, H.-T.; Nou, I.-S. Identification of nbs-encoding genes linked to black rot resistance in cabbage (Brassica oleracea var. capitata). Mol. Biol. Rep. 2018, 45, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Poczai, P.; Varga, I.; Laos, M.; Cseh, A.; Bell, N.; Valkonen, J.P.; Hyvönen, J. Advances in plant gene-targeted and functional markers: A review. Plant Methods 2013, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Lin, Y.; Chen, G.; Shen, Y.; Liu, J.; Zhang, S. Genome-scale identification of resistance gene analogs and the development of their intron length polymorphism markers in maize. Mol. Breed. 2012, 29, 437–447. [Google Scholar] [CrossRef]
- Slabaugh, M.B.; Yu, J.K.; Tang, S.; Heesacker, A.; Hu, X.; Lu, G.; Bidney, D.; Han, F.; Knapp, S.J. Haplotyping and mapping a large cluster of downy mildew resistance gene candidates in sunflower using multilocus intron fragment length polymorphisms. Plant Biotechnol. J. 2003, 1, 167–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockah-Shmuel, L.; Tóth-Petróczy, Á.; Sela, A.; Wurtzel, O.; Sorek, R.; Tawfik, D.S. Correlated occurrence and bypass of frame-shifting insertion-deletions (InDels) to give functional proteins. PLoS Genet. 2013, 9, e1003882. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, Y.; Zhai, W.; Deng, J.; Wang, H.; Cui, Y.; Cheng, F.; Wang, X.; Wu, J. Development of indel markers for Brassica rapa based on whole-genome re-sequencing. Theor. Appl. Genet. 2013, 126, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Yang, M.; Liu, H.; Tao, Y.; Mei, J.; Zhao, Y. Genetic analysis and molecular characterization of chinese sesame (Sesamum indicum L.) cultivars using insertion-deletion (InDel) and simple sequence repeat (SSR) markers. BMC Genet. 2014, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J. High-throughput SNP genotyping to accelerate crop improvement. Plant Breed. Biotechnol. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Liu, R.H.; Meng, J.L. Mapdraw: A microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi Chuan 2003, 25, 317–321. [Google Scholar] [PubMed]
- Soderlund, C.; Bomhoff, M.; Nelson, W.M. Symap v3. 4: A turnkey synteny system with application to plant genomes. Nucleic Acids Res. 2011, 39, e68. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.I.; Schein, J.E.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crosses | Generation | Susceptible (PDI > 20) | Resistant (PDI ≤ 20) | Expected Ratio (S:R) | Chi-Square (χ2) | p |
|---|---|---|---|---|---|---|
| PI482399 | P1 | 0 | 15 | |||
| Cornell ZPPM 339 | P2 | 15 | 0 | |||
| Cornell × PI482399 | F1 | 15 | 0 | |||
| Cornell × PI482399 | F2 | 126 | 42 | 3:1 | 0.911 | 0.639 |
| Sl. No. | Gene ID | Domain | Primer (5’–3’) | Product Length (bp) |
|---|---|---|---|---|
| 1 | MELO3C022157 | TIR-NBS-LRR | F: ATGGTGCTTGAGAGAATTGG | 876 |
| R: GTTCATGATTGGGGTGAGAA | ||||
| 2 | MELO3C022154 | TIR-NBS-LRR | F: CTTTCGAGGCGAAGATACAC | 1695 |
| R: CAGCCTAAGTTGGTGCATTC | ||||
| 3 | MELO3C022152 | TIR-NBS-LRR | F: GGCTTCTCCAGCAACAATAA | 1703 |
| R: ATTCCCACATGCCAAGTTT | ||||
| 4 | MELO3C022148 | TIR-NBS-LRR | F: GTTGGCAATCTCTCTGGATG | 1009 |
| R: GCATTTGTATCTTCTCATGTGG | ||||
| 5 | MELO3C022146 | TIR-NBS-LRR | F: TTTTAGAGGCGAAGATACTCGT | 1215 |
| R: CAAAGCTTGTGGATGTCCTT | ||||
| 6 | MELO3C022145 | TIR-NBS-LRR | F: CCACCATCTCTGCTTGAGTT | 1915 |
| R: GTGGAGAAATCATCCACGAC | ||||
| 7 | MELO3C022144 | TIR-NBS-LRR | F: TCATCGTCTTCTTTGGATCG | 1427 |
| R: GGACTATAACCAAAACTCTCCA | ||||
| 8 | MELO3C022143 | TIR-NBS-LRR | F: GATGCTGCTGATGGACTTCT | 1648 |
| R: CATTCTCCCAAGCTGTGC | ||||
| 9 | MELO3C021485 | RLK | F: CCTGAAGATGATATAAGGTGTC | 1696 |
| R: GTGGGAATTCATCATCTGGTTC | ||||
| 10 | MELO3C025516 | TIR-NBS-LRR | F: AGTCGAGTCATCGTTACAACA | 849 |
| R: CTCAAGCTCCAAGAGGTTTGTG | ||||
| 11 | MELO3C002877 | TIR-NBS-LRR | F: GGCCAAAGAGTTTAGGAATATC | 933 |
| R: ATGATGATTCGACTCCCTGG | ||||
| 12 | MELO3C002928 | LRR | F: GCACCAATTTCCATTCATACCC | 796 |
| R: GCCAGGTGGTATTATCCCAGAC | ||||
| 13 | MELO3C005099 | RLK | F: TGGTGTTATGGCGGAGGGTTC | 775 |
| R: ATTGAGAACTCTAAGCGAGC | ||||
| 14 | MELO3C005209 | LRR | F: GATGCAAAATGCCACCCTGA | 949 |
| R: TGGGCATAATCTCTCCACGGA | ||||
| 15 | MELO3C005236 | RLK | F: AGTTGTAGGTAGTGTGCCTG | 860 |
| R: GATTCTGAAGCCTTCCAAGC | ||||
| 16 | MELO3C005267 | RLK | F: GCATGTGAAATGTGATTCTCAG | 822 |
| R: GCATTCACTCGAGGATCAC | ||||
| 17 | MELO3C005339 | RLK | F: CCTTGTTCTTGGAATGGGGTTG | 999 |
| R: ATCATACTGCTCCACATTCTG | ||||
| 18 | MELO3C005374 | RLK | F: CTCTCTGTCAAGTTTCAGCC | 1000 |
| R: AGATTATCACCCACCCATGTAG | ||||
| 19 | MELO3C005413 | RLK | F: GAAGGCGGCTTTGGACCCTGA | 905 |
| R: TTCGACGACGGTATTGCGTG | ||||
| 20 | MELO3C005427 | LRR | F: ATGATACTGCTTCTTCTACACC | 1242 |
| R: CTGTGAATGTGGAAAGGTCCCA | ||||
| 21 | MELO3C005450 | LRR | F: GTCCATAGCAGCAGCAGCTTGC | 865 |
| R: TCCAAACACATCTCCACCAAG | ||||
| 22 | MELO3C005504 | LRR | F: ATGGACTCAGAGGCAGGGC | 889 |
| R: GGTCATCAATCTTCTCCAGC | ||||
| 23 | MELO3C005660 | RLK | F: AGCCGACAAGAACTTGAATTAGC | 721 |
| R: TGTACGACATCACTTCTGGCA | ||||
| 24 | MELO3C005864 | RLK | F: ATGTCAGAGCCGATCAAAGAC | 734 |
| R: GCTAACCCCATTGCAGAACCTCC | ||||
| 25 | MELO3C005732 | RLK | F: CCTCACTCTCGCTCTCTTAATTGG | 846 |
| R: GTCAAAAGCATAATGGCACCC | ||||
| 26 | MELO3C025088 | RLK | F: ATCGGCCGCCAAAATCTCCAAG | 791 |
| R: GAAAGCAATGTATTCACTCGTG | ||||
| 27 | MELO3C025083 | RLK | F: CGGAATCACCACCGGCTCTGTT | 1063 |
| R: CATCTCTTAACCTCTTAACCAC | ||||
| 28 | MELO3C005170 | RLK | F: CAACAAGCAAAGACGATTCGAC | 1010 |
| R: GAGAAGGTTCCTATGCACTGCC | ||||
| 29 | MELO3C005428 | LRR | F: TCCATAACTGCAGCAGCTTGC | 931 |
| R: CCAAGTGGTTTCCGGAGAGGTC | ||||
| 30 | MELO3C005435 | LRR | F: GACCAACCAAATTAAACACCCA | 922 |
| R: ACCTGGCGGCAAAGGACCCGT | ||||
| 31 | MELO3C022083 | LRR | F: ATGGGAATGAAGAGAACTGAGG | 520 |
| R: GCTGTGTCATGTTAGCTAAAGAC | ||||
| 32 | MELO3C022040 | RLK | F: TCTCAAACACCATCAACGAAGC | 1066 |
| R: GGCAAATCCCACGCTACTACAC | ||||
| 33 | MELO3C005451 | LRR | F: ATGATGTTGCTGCTTCTCCCAT | 874 |
| R: GTGGAACTATAATCTTGGTGGAG | ||||
| 34 | MELO3C025517 | TIR-NBS-LRR | F: GCATCCTTATCTTCTCCTCCGC | 557 |
| R: GAAGCCGGAGAGAGTGAAAACA | ||||
| 35 | MELO3C005508 | LRR | F: ATGGCTTCAACGGAAATCTCAAC | 1012 |
| R: ACCACTCCACAACTTTGACAGT |
| Gene ID | InDel Marker | Primer (5’–3’) | Product Size | |
|---|---|---|---|---|
| MELO3C022157 | Kh-GSB9-1 | F | GTTAGGAAACAACAGACCTCCA | 266 |
| R | CAGAACGCACAAAACTCAAAGGAC | |||
| MELO3C022157 | Kh-GSB9-2 | F | CCTAATAGTCCTTTGAGTTTTGTGCG | 409 |
| R | GGTGTGCTTGGATTGGCTTTCT | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.Z.; Rahim, M.A.; Natarajan, S.; Robin, A.H.K.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Gummy Stem Blight Resistance in Melon: Inheritance Pattern and Development of Molecular Markers. Int. J. Mol. Sci. 2018, 19, 2914. https://doi.org/10.3390/ijms19102914
Hassan MZ, Rahim MA, Natarajan S, Robin AHK, Kim H-T, Park J-I, Nou I-S. Gummy Stem Blight Resistance in Melon: Inheritance Pattern and Development of Molecular Markers. International Journal of Molecular Sciences. 2018; 19(10):2914. https://doi.org/10.3390/ijms19102914
Chicago/Turabian StyleHassan, Md Zahid, Md Abdur Rahim, Sathishkumar Natarajan, Arif Hasan Khan Robin, Hoy-Taek Kim, Jong-In Park, and Ill-Sup Nou. 2018. "Gummy Stem Blight Resistance in Melon: Inheritance Pattern and Development of Molecular Markers" International Journal of Molecular Sciences 19, no. 10: 2914. https://doi.org/10.3390/ijms19102914