Field effects of oxygenated monoterpenes and estragole combined with pheromone on attraction of Ips typographus and its natural enemies
 Antonioni Acacio Campos Moliterno1†
Antonioni Acacio Campos Moliterno1†  Rastislav Jakuš1,2
Rastislav Jakuš1,2  Roman Modlinger1
Roman Modlinger1  C. Rikard Unelius3
C. Rikard Unelius3  Fredrik Schlyter1,4
Fredrik Schlyter1,4  Anna Jirošová1*†
Anna Jirošová1*†- 1Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Prague, Czechia
- 2Institute of Forest Ecology, Slovak Academy of Sciences, Zvolen, Slovakia
- 3Faculty of Health and Life Sciences, Linnaeus University, Kalmar, Sweden
- 4Department of Plant Protection Biology, Chemical Ecology, Swedish University of Agricultural Sciences, Alnarp, Sweden
Introduction: Central European Norway spruce monocultures face Ips typographus outbreaks due to decreasing resistance. These beetles use volatile compounds to communicate and select suitable host trees. Spruce trees, beetles, and their symbiotic ophiostomatoid fungi emit oxygenated monoterpenes, including 1,8-cineole, α-terpineol, camphor, carvone, terpinen-4-ol, isopinocamphone, and pinocamphone, and the phenylpropanoid estragole, particularly in the infestation phase. These compounds trigger strong responses in I. typographus antennae, motivating our field study.
Objective: This study aimed to assess how adding these compounds to the aggregation pheromone of Ips typographus modulates the attraction of this bark beetle and its natural enemies.
Methods: In combination with I. typographus pheromone, estragole, 1,8-cineole, (±)-camphor, (–)-carvone, alpha-terpineol, (–)-terpinen-4-ol, (+)-pinocamphone, and (+)-isopinocamphone at low, medium, and high doses were tested in pheromone traps at two sites in the Czech Republic.
Results: All 1,8-cineole doses and the high estragole dose acted as anti-attractants for I. typographus, whereas all (+)-isopinocamphone doses enhanced their attraction to pheromone. Catches of natural enemies, the Staphylinidae and Pteromalidae, varied by location.
Conclusion: 1,8-cineole, isopinocamphone, and estragole may play vital roles in tritrophic interactions among spruce trees, and I. typographus and its natural enemies, and these compounds may be developed into new or enhanced semiochemical-based pest control methods.
1 Introduction
In Central Europe, Norway spruce (Picea abies) (L.) Karst. (Pinales: Pinaceae) has been severely affected by infestations of the spruce bark beetle, Ips typographus (L.) (Coleoptera: Curculionidae: Scolytinae), that in the Czech Republic have resulted in timber losses of 5.9 mil. m3 in 2017 and 26.2 mil. m3 in 2020 (Hlásny et al., 2022). In outbreak regions, managing bark beetles often involves applying insecticides to P. abies trunks or stored timber to eliminate the emerging beetles (Fettig and Hilszczański, 2015). However, the use of pesticides can negatively impact the forest ecosystem, including beneficial bark beetle predator species (Hlásny et al., 2019). The development of alternative, eco-friendly strategies in forestry is a logical progression. One such strategy involves utilizing semiochemicals, compounds that mediate the interactions of beetles with each other and other organisms. These signals enable beetles to locate a mate or host tree by providing intraspecific and interspecific chemical information (Bergström, 2007). Aggregation pheromone components, produced by male beetles after successful colonization (Birgersson et al., 1984; Ramakrishnan et al., 2022), have been employed for monitoring and controlling I. typographus populations (Heber et al., 2021). Furthermore, recent research has focused on the management potential of kairomones, compounds originating from both host spruce trees and non-host trees, e.g., broadleaf trees (Zhang and Schlyter, 2004; Jakuš et al., 2022). Bark beetles possess a sophisticated olfactory system that enables them to detect and distinguish the chemical composition and quantity of these odors (Andersson, 2012).
The principal olfactory stimulants for I. typographus emitted by P. abies are primarily composed of high-abundance monoterpenes such as α-pinene (23–39%), β-pinene (25–58%), β-phellandrene (5–19%), limonene (1.5–4%), myrcene (1.6–3.4%), Δ-3-carene (0.6–1.1%), and camphene (0.2–1.1%; Netherer et al., 2021). However, recent comparative analysis utilizing I. typographus antennae as biological detectors (gas chromatography coupled with electroantennography, GC-EAD) has identified several novel compounds present in relatively small amounts but exhibiting high activity with the beetles' antennae (Kalinová et al., 2014; Schiebe et al., 2019). These compounds include oxygenated monoterpenes, 1,8-cineole (eucalyptol), trans-4-thujanol (sabinene hydrate), camphor, pinocarvone, pinocamphone, isopinocamphone, terpinen-4-ol, alpha-terpineol, carvone, and phenylpropanoid estragole (4-allylanisole and methyl chavicol). In single-cell electrophysiological studies, researchers identified 24 classes of olfactory sensory neurons (OSN) within olfactory sensillae for I. typographus (Hallberg, 1982). Plant odor-responding OSNs exhibit a variety of response specificities from broadly tuned OSNs for host monoterpene hydrocarbons to several highly specific OSN classes responding mainly to oxygenated monoterpenes (1,8-cineole, isopinocamphone, trans-4-thujanol, or verbenone; Andersson et al., 2009; Schiebe et al., 2019; Kandasamy et al., 2023).
In Norway spruce, oxygenated monoterpenes are minor compounds (~1% representation), and their content is influenced by tree health and stress levels (Netherer et al., 2021). The production of oxygenated monoterpenes in trees naturally occurs through the cytochrome P450-catalyzed oxidation of monoterpene hydrocarbons or by cyclization of oxygenated intermediates (Celedon and Bohlmann, 2019). The release rate of oxygenated monoterpenes, including 1,8-cineol, camphor, pinocarvone, terpinen-4-ol, and α-terpineol, from healthy trees at 24°C ranges from 0.1 to 7 μg/m2/h of stem surface area (Ghimire et al., 2016). In infested trees, these rates increased 10–100 times (Jaakkola et al., 2022), and in cut trees, they increased 10 times (Schiebe et al., 2019).
Bark beetle symbiotic ophiostomatoid fungi generate oxygenated terpenes in laboratory conditions when they are inoculated onto a wood substrate (Kandasamy et al., 2023). In the forest, fungi may assist beetles in colonizing healthy trees by being involved in detoxifying host defense terpenes (Krokene, 2015; Kandasamy et al., 2021). Additionally, the beetles themselves generate oxygenated monoterpenes, as they metabolize toxic terpenes while feeding on the spruce tree's phloem (Blomquist et al., 2021). The detoxification process of terpenes involves a series of steps. In the first step, a hydroxyl group is introduced to a terpene molecule by cytochrome P450 catalysis. This modification increases the molecule's polarity and solubility in water, enabling beetles to eliminate it (Blomquist et al., 2021). In subsequent steps, the resulting terpenic alcohols are either excreted from the body or bound to detoxification conjugative molecules, such as fatty acid esters (Chiu et al., 2018) or glycosylates (Dai et al., 2021). This mechanism was studied in Dendroctonus ponderosae (Chiu, 2018; Chiu et al., 2019), Dendroctonus armandi (Dai et al., 2021), as well as in Ips species (Blomquist et al., 2021; Ramakrishnan et al., 2022) that feed on conifer trees, as these trees possess terpenes as a defense trait. This adaptation allows the beetles to overcome the tree's defenses and successfully colonize it. It is theorized that during evolution, some of these detoxification products, such as cis-verbenol in I. typographus, started to serve as aggregation pheromones for the bark beetles (Blomquist et al., 2021; Schebeck et al., 2023).
The exact behavioral role of all host-produced oxygenated monoterpene semiochemicals in bark beetles is not fully understood. However, according to the primary attraction theory proposed by Lehmanski et al. (2023), these compounds may play a role in helping male bark beetles detect weakened host trees, thereby facilitating successful colonization. Other compounds, e.g., 1,8-cineole and trans-4-thujanol, have proved to be potent anti-attractive compounds inhibiting beetle attraction to their pheromone (Jirošová et al., 2022b). 1,8-cineole has been identified as a potential predictor of bark beetle-resistant trees, along with several other specialized metabolites (Schiebe et al., 2012). Higher levels of trans-4-thujanol were detected in younger Norway spruces. Given that this compound has demonstrated repellency in high doses in laboratory olfactometer studies, it provides a potential explanation for the reduced attraction of I. typographus to trees below the age of 60 years (BlaŽyte-Cereškiene et al., 2016). The activity of 1,8-cineole and trans-4-thujanol for I. typographus has been evaluated in a field trapping experiment using different doses in combination with pheromones. Both compounds demonstrated a similar level of dose-dependent anti-attractant activity, with trans-4-thujanol inhibiting more the captures of females than males (Jirošová et al., 2022b). These compounds have been tested in combination with other anti-attractants for the protection of spruce trees in various forest environments, such as fresh forest edges or fragmented forests (Jakuš et al., 2022; Jirošová et al., 2022a). Furthermore, individual compounds trans-4-thujanol, (+)-isopinocamphone, camphor, and terpinen-4-ol were tested in different doses in two-choice Petri dish walking laboratory tests to assess their attractiveness to I. typographus. The activity of these compounds was largely insignificant, and only trans-4-thujanol and camphor at a high dose were attractive. In contrast, a more complex mixture of oxygenated monoterpenes, generated through the metabolization of (–)-β-pinene and (–)-bornyl acetate by the ophiostomatoid fungus Grosmania polonica, exhibited dose-dependent attractivity in the test (Kandasamy et al., 2023).
In multi-trophic interactions, the bark beetle predator-prey relationships are influenced by qualitative and quantitative aspects of host tree compounds and prey pheromones (Erbilgin and Raffa, 2001; Netherer et al., 2021). The natural enemies associated with I. typographus include Hymenoptera: Pteromalidae (Rhopalicus spp.), Diptera: Dolichopodidae (Medetera spp.), Coleoptera: Cleridae (Thanasimus spp.), and Coleoptera: Staphylinidae (Nudobius sp. and Quedius sp.; Wegensteiner et al., 2015). Laboratory bioassays demonstrated the strong attraction of female Rhopalicus spp. to infested logs, with antennal responses to oxygenated monoterpenes including pinocamphone, pinocarvone, and the phenylpropanoid estragole (Pettersson, 2001; Pettersson and Boland, 2003). Similarly, camphor, pinocamphone, and terpinen-4-ol derived from bark beetle-associated microorganisms in infested spruce logs exhibited strong attraction with Medetera signaticornis Loew (Sousa et al., 2023). Staphylinidae feeds on a wide range of prey, and some species are hypothesized to be associated with bark beetles (Pelto-Arvo, 2020). However, their specific association with host tree volatiles, such as oxygenated monoterpenes and I. typographus pheromone, remains unexplored.
The foraging strategies of some I. typographus predators, Medetera spp. and Thanasimus spp., could be governed by a more complete blend of the aggregation pheromone and host volatiles (represented by oxygenated monoterpenes; Hulcr et al., 2006). The hemiterpene pheromone component 2-methyl-3-buten-2-ol by itself does not increase predator catch, while the minor component ipsdienol does (Bakke and Kvamme, 1981; Hulcr et al., 2006; Raffa, 2014).
Further research is necessary to achieve a more comprehensive understanding of the mechanisms and functions of oxygenated monoterpenes and estragole in the behavior of bark beetles and their predators.
In this study, we aimed to address the following specific questions:
1. Can individual host tree-oxygenated monoterpenes and estragole enhance or reduce the attractiveness of I. typographus pheromones in field traps for capturing bark beetles?
2. If so, which of the three tested decadic steps in release rates, namely, low (representing conditions closest to natural levels), medium, and high (representing doses used in management), exhibits the highest efficacy?
3. What is the species composition and abundance of predatory insects attracted to the tested compounds using the methodology described?
4. Can we propose a specific role for the tested compounds in bark beetle ecology?
We conducted a field trapping experiment to investigate the activity of eight synthetic host tree compounds (estragole, 1,8-cineole, (±)-camphor, (–)-carvone, (–)-α-terpineol, (–)-terpinen-4-ol, (+)-pinocamphone, and (+)-isopinocamphone) for I. typographus and some of its natural enemies. The compounds were tested at low, medium, and high release rates, in combination with components in pheromone barrier traps.
2 Materials and methods
2.1 Experimental area and design
For field experiments, we chose two locations in the Czech Republic. The first location was in North Moravia in Libavá (Military Forests, latitude 49°38′49 “N, longitude 017°33′50” E, 350 m above sea level). It consisted of a 40-year-old Norway spruce forest that has been heavily impacted by a bark beetle outbreak since 2015 (Brázdil et al., 2022). The experiment in Moravia was conducted from 18 May to 3 June 2022. The second location, Kostelec nad Cernými Lesy (Forests CZU; latitude: 49°55′57 “N, longitude: 014°55′13” E, 600 m above sea level), consisted of a 70–90-year-old Norway spruce forest. Traps were placed in a 2-year-old clearing measuring ~200 m × 300 m. The experiment was carried out from 3 June to 28 July 2022. The experiment was designed identically in both locations (Figure 1).
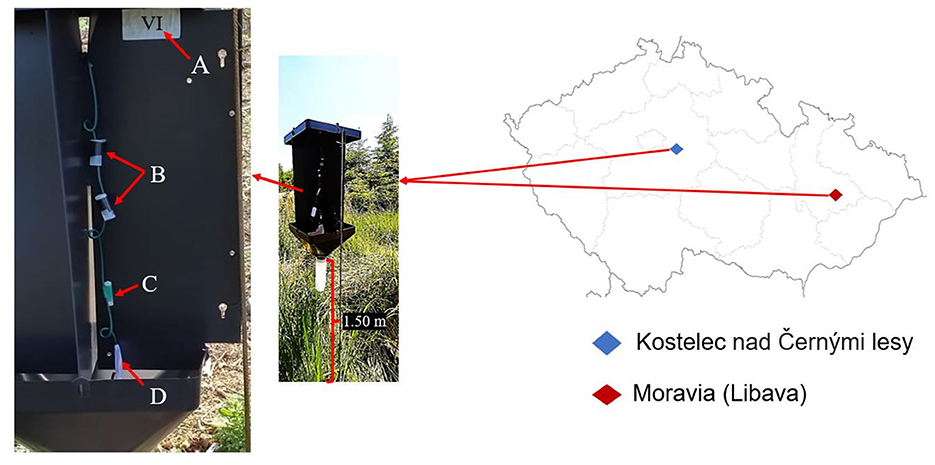
Figure 1. Field experiment sites and cross-panel field traps in the Czech Republic: (A) Trap label, (B) synthetic aggregation pheromone dispensers, (C) synthetic host tree odor dispenser, and (D) bait label for treatment identification.
The activity of estragole, 1,8-cineole, (±)-camphor, (–)-carvone, (-)-α-terpineol, (–)-terpinen-4-ol, (+)-pinocamphone, and (+)-isopinocamphone was tested at three different doses, represented by their release rates evaporated/sublimated from the dispenser (nominal 0.1, 1, and 10 mg/day, Table 1). The doses were determined based on the published releases of oxygenated monoterpenes from healthy trees at 24°C, which varied from 0.1 to 7 μg/m2/h of stem surface (Ghimire et al., 2016). When considering a tree stem with a 50 cm diameter and an exposed surface area of ~24 m2 (representing 15 m of stem height vulnerable to bark beetle attack), the estimated daily release rate of these oxygenated monoterpenes over a 24-h period would be ~0.5–4 mg/day. We used the pure enantiomers of (–)-carvone, (–)-terpinen-4-ol, (+)-pinocamphone, and (+)-isopinocamphone, which triggered a higher response on the I. typographus antennae (Schiebe et al., 2012; Hou et al., 2021; Kandasamy et al., 2023) and were commercially available or synthesized in the laboratory (Ganji et al., 2020). Experimental dispensers were designed in the laboratory, and their exact laboratory and field release rates were established using the gravimetric method and measured six times (Jirošová et al., 2022b) in a laboratory fume hood (temperature 25°C and airflow 0.5 m/s) and in the field under the same weather conditions as the experiments (Table 1).
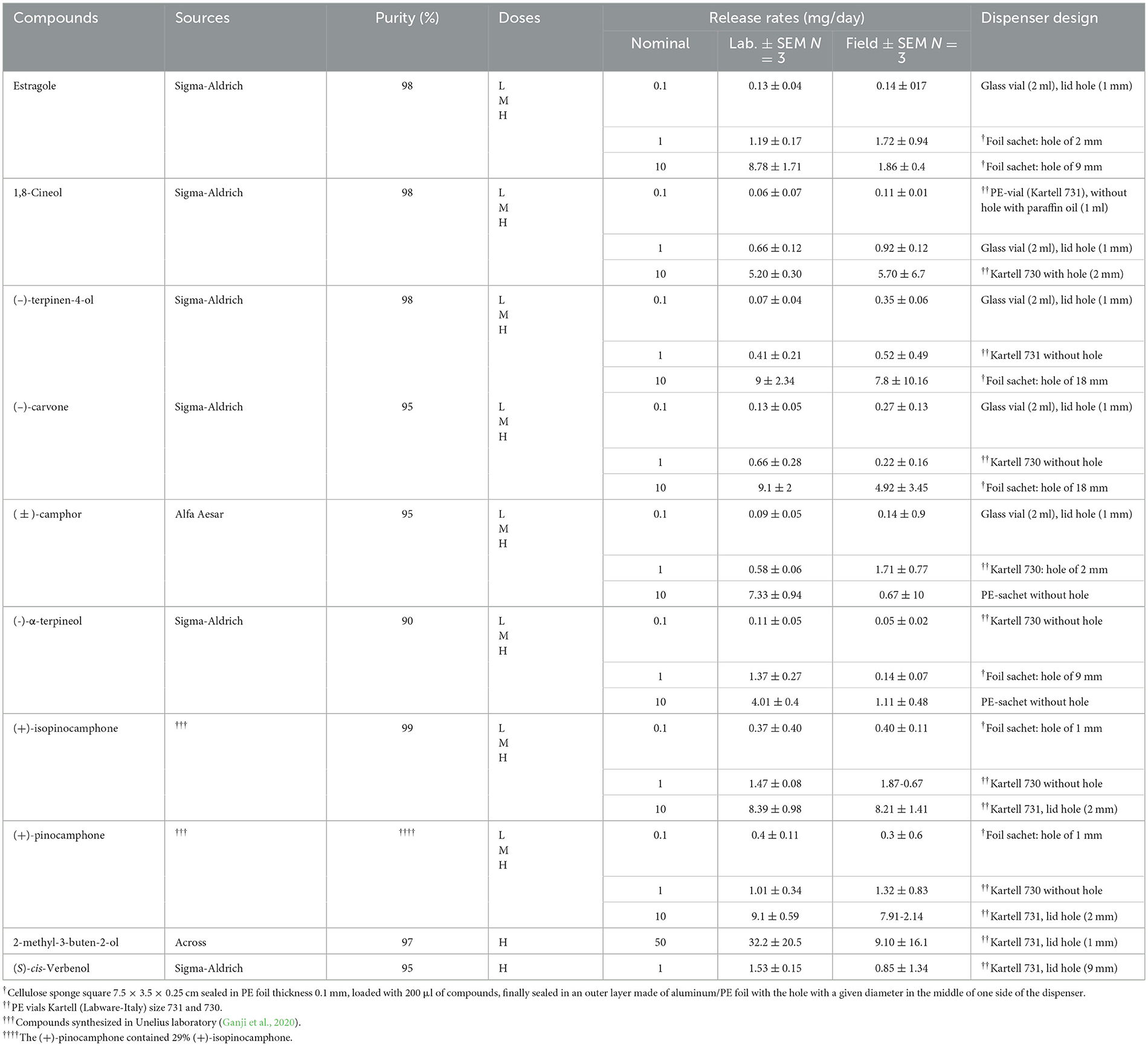
Table 1. The gravimetric establishment of released rates of tested compounds performed in the laboratory and field.
In the field, the intercept pheromone traps (Ecotrap/Fytofarm, Ltd., Bratislava; Slovakia) were mounted on poles 1.5 m above the ground in rows >30 m from any forest edge. The distance between traps was >15 m (Supplementary Figure 1). In each field location, 32 intercept pheromone traps were baited with dispensers with I. typographus pheromone (2-methyl-3-buten-2-ol at 9.1 mg/day and (S)-cis-verbenol at 0.9 mg/day). In 24 of these traps, an additional dispenser was placed with one of the eight test compounds in one of three doses.
For each compound, one block represented four traps arranged in a row: three traps with different doses in combination with pheromone and one trap with pheromone-only (Control). The position of the tested baits among these four traps was changed four times according to the randomization scheme in a Latin square design (Evans et al., 2020). These four rotations were repeated twice for each compound, resulting in a total of eight collections of catches for each treatment. Insects collected during the field experiment in both localities were preserved in ethanol for further analysis. Predators and parasitoids were sorted by family and identified at the genus level. The identification of Pteromalidae wasps followed the methods described by Peck et al. (1964), Graham (1969), and Bouček and Rasplus (1991). For Staphylinidae (rove beetles), the identification followed the guidelines provided by Arnett and Thomas (2000) and Navarrete and Newton (2003).
2.2 Statistical analysis
For the evaluation of the effects of each individual compound, a separate regression model was fitted with the relative number of I. typographus as the dependent variable. The relative number was expressed as the number of insects of a single taxon captured by a treatment within a block divided by the total catches by the block for a single catch collection. Due to the experimental design, we utilized a mixed-effects model approach (Zuur et al., 2009). The random part of the model was, in all cases, trap-nested in the locality. During model building and validation, an appropriate distribution function was selected by minimizing the Akaike information criterion (AIC), and the significance of the model was tested by the likelihood ratio test (χ2). Between the best models, only two distribution functions were selected (Table 2): generalized poison distribution (Joe and Zhu, 2005) and negative binomial distribution in quadratic parameterization, according to Hardin and Hilbe (2007). We used a t-test to compare the response to each compound dose against the pheromone-only control. The model formulation was performed in R version 4.3.1 (R Core Team, 2023) in the package glmmTMB following the procedures described by Brooks et al. (2017).
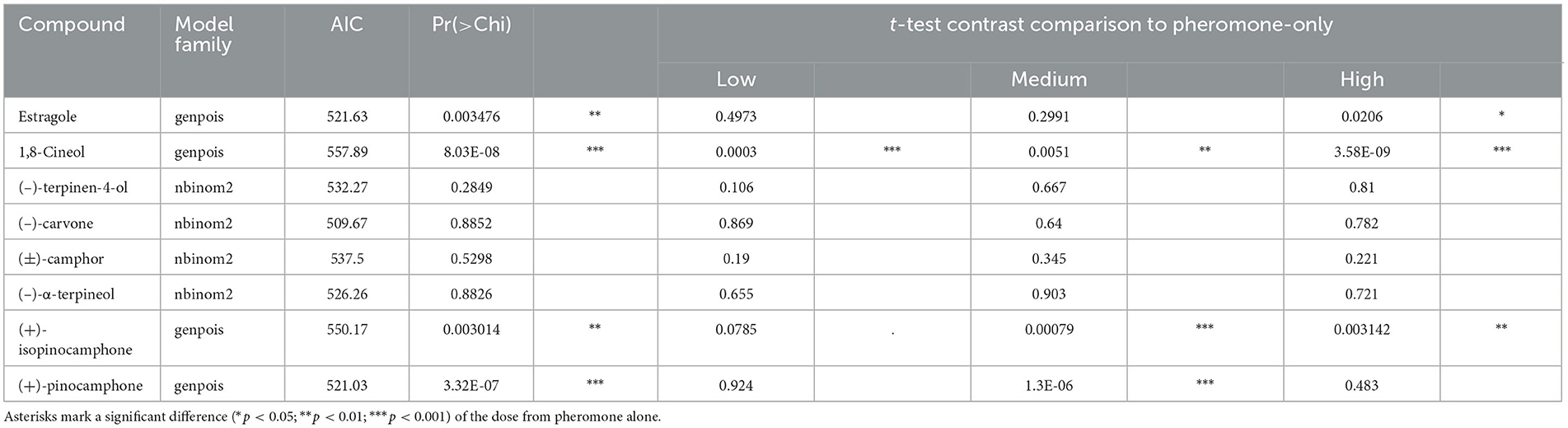
Table 2. Testing of relative catches of I. typographus in the tested compounds in different doses.
3 Results
3.1 Ips typographus response to tested compounds in combination with aggregation pheromone
During field experiments, a total of 39,650 I. typographus adults were caught. The number of adults captured in Kostelec (N = 28,931) was 2.7 times higher than in Moravia (N = 10,719). However, the pattern of catches for the tested compounds was almost the same. Compounds with significantly different catches in the treatments: estragole (χ2; p < 0.01), 1,8-cineole (χ2; p < 0.001), (+)-pinocamphone (χ2; p < 0.01), and (+)-isopinocamphone (χ2; p < 0.01; Table 1, Figure 2). For 1,8-cineol, all three doses in combination with pheromones resulted in significantly fewer beetles caught than the pheromone-only control, with stronger effects observed for the high dose (t-test; p < 0.001, p < 0.01, p < 0.001; Table 2, Figure 2). For estragole, the inhibitory effect was significant only for the high dose (t-test; p < 0.05), while for pinocamphone, it was observed at the medium dose (t-test; p < 0.001). However, regarding the medium dose of pinocamphone, there were different catch rates in Libava and Kostelec (Figure 2), suggesting a potential problem with the dispenser used in Kostelec. Conversely, isopinocamphone resulted in statistically higher catch rates at both high and medium doses (t-test; p < 0.01 and p < 0.001, respectively) compared to the pheromone-only treatment (Figure 2). A low dose of isopinocamphone showed the same nearly significant trend (t-test; p = 0.08; Table 2). Catches of beetles of remaining tested compounds in combination with pheromone, including (±)-camphor, (–)-carvone, (-)-α-terpineol, (-)-terpinen-4-ol, and pheromone-only, did not exhibit significant differences (Table 2). Additionally, there were no significant differences in catches between the individual doses and the pheromone-only control group for any of these compounds (Supplementary Figure 2).
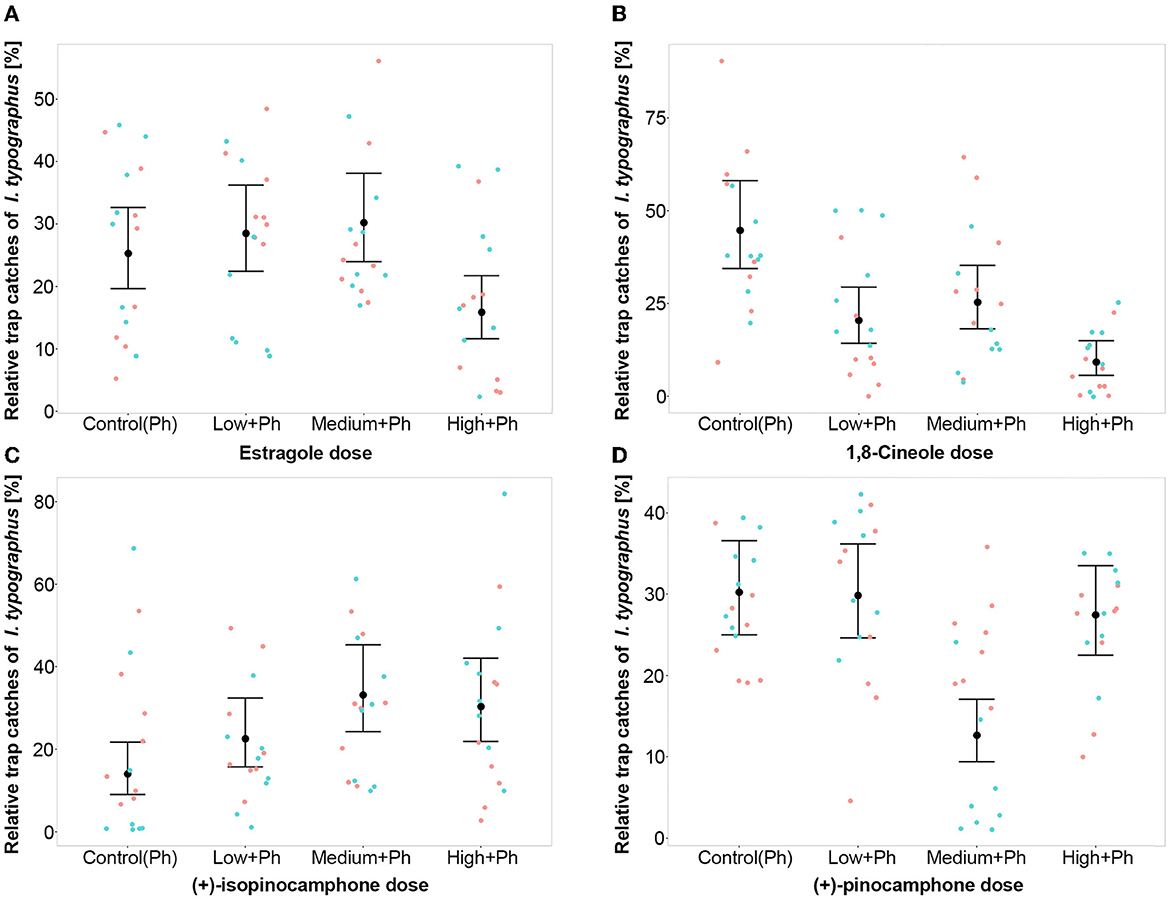
Figure 2. Relative number of I. typographus captured in traps baited with pheromone attractant (Ph) alone or attractant with host volatiles (A) estragole, (B) 1,8-cineole, (C) (+)-pinocamphone, and (D) (+)-isopinocamphone in a low-medium, high-release dose combined with pheromone (Ph) and control pheromone-only. The bigger black dot in the middle of the vertical line is a posterior mean value and whiskers. Error bars represent the 95% confidence interval, and colored points represent the raw catches. The smaller dots are original values: blue Kostelec and red Libavá.
For each compound, a separate model (GLMM) was created with the formula: Relative count of I. typographus ~ Compound dose + (1|Locality). The appropriate distribution function (model family: genpois—Generalized poison; nbinom2—Negative binomial), Akaike information criterion (AIC), and significance test (χ2) are stated for each compound. The results of the t-test contrast comparison given for each compound and dose combination against pheromone-alone (control). Asterisks mark a significant difference (p < 0.05 *; p < 0.01 **; p < 0.001 ***) of the dose from pheromone alone.
3.2 Predatory insect response to tested compounds in combination with aggregation pheromone
The catch of natural enemies was three times higher in Kostelec compared to Moravia, which corresponds to a higher number of bark beetle catches in Kostelec. Four families of natural enemies of bark beetles were identified with a prevalence of Pteromalidae and Staphylinidae (Supplementary Tables 1, 2). In Kostelec, there were 93 specimens of parasitoid wasps belonging to the genus Rhopalicus sp. (Hymenoptera: Pteromalidae), while in Moravia, there were only 13 specimens. Additionally, in Kostelec, 19 specimens of rove beetles (Coleoptera: Staphylinidae) were caught, and in Moravia, 23 specimens (Table 3).

Table 3. Natural enemies of bark beetles (Staphylinidae and Pteromalidae) captured in synthetic host tree compounds: estragole, 1,8 cineole, camphor, carvone, alpha-terpineol, terpinene-4-ol, (+)-pinocamphone, and (+)-isopinocamphone, in combination with pheromone and pheromone-only controls.
The ratio between these two groups appeared different between the two locations, with Kostelec having a higher proportion of Pteromalid wasps compared to Staphylinids, while in Moravia, we observed the opposite trend. There were a few catches recorded for Medetera sp. (Diptera: Dolichopodidae), with five specimens in Kostelec and three specimens in Moravia. There were four catches of Lonchaea sp. (Diptera: Lonchaeidae), three in Kostelec, and one in Moravia. For Thanasimus sp. (Coleoptera: Cleridae), there were two catches in Kostelec and three catches in Moravia.
The catches of Rhopalicus sp. wasps did not show significant differences between compounds and their doses. The number of caught specimens in Moravia and Kostelec is listed in the Supplementary Tables 1, 2.
4 Discussion
4.1 Response of Ips typographus to tested compounds
The variability in total catches of I. typographus and its predators, as well as their varying distributions across different treatments in the experimental locations of Kostelec and Moravia, could be attributed to the unique weather conditions experienced during each field experiment, as described in Supplementary Figure 3. Furthermore, this variation may have been influenced by the season impacting the flight activity of the beetles. The variation between catches in blocks of traps may have been due to different locations of blocks within the clearing, in terms of their distance to the forest edge and wind speed and direction. The trap catches could also have been influenced by different ages of spruce stands near the experiments.
In many cases, the beetles exhibited weak differences in response to the tested compounds in combination with the pheromone compared to the pheromone alone, and the observed effects were only marginally significant. However, there was a noticeable anti-attractive effect observed with 1,8-cineole, a compound for which similar findings and similar trends have been reported previously (Andersson et al., 2010; Binyameen et al., 2014; Jirošová et al., 2022b).
New findings were the anti-attractive effect of a high dose of estragole and the effect of (+)-isopinocamphone on enhancing the attractive activity of the pheromone for I. typographus. There was also a less clear inhibition effect of a medium dose of pinocamphone, probably caused by a defective test dispenser for a medium dose at Kostelec. In contrast, when α-terpineol, camphor, carvone, or terpinen-4-ol were added to the pheromone, they did not alter its attractiveness, despite these compounds eliciting strong responses from I. typographus antennae (Kalinová et al., 2014; Schiebe et al., 2019).
The ecological role of estragole, 1,8-cineole, and isopinocamphone in the interaction between I. typographus and Norway spruce trees was investigated by Schiebe (2012, 2019). The amount of these compounds, along with other oxygenated monoterpenes found in spruce, increased in felled trees and in standing trees after the application of the plant hormone analogue methyl jasmonate. The quantity of these compounds was negatively correlated with the density of bark beetle attacks when the beetles infested the felled trees, and the standing trees that exhibited a higher induction of these compounds were able to survive a natural bark beetle infestation.
The effect of estragole has been tested on several bark beetle species (Coleoptera: Curculionidae: Scolytinae), including Dendroctonus brevicomis LeConte (Hayes and Strom, 1994), Ips pini (Say), Ips latidens (Leconte) (Joseph et al., 2001), and Tomicus piniperda (Curculionidae: Scolytinae) (Haack et al., 2004). Its inclusion in their host odor blend resulted in reduced trap catches. Estragole was also reported to interrupt the responses of bark beetle species Dedroctonus simplex (Le Conte) and D. ruffipenis (Kirby) to their attractive pheromone components (Werner, 1995). However, recent research has revealed that the addition of estragole increased catches of both Dendroctonus frontalis (Zimmermann) and D. terebrans (Olivier) on their pheromone lures (Munro et al., 2020). Based on these findings, we suggest that this semiochemical has variable ecological roles for these different species.
In our study, (+)-isopinocamphone caused a synergistic increase in beetle catches when added to the pheromone. This observation, combined with the fact that bark beetles possess specialized sensilla on their antennae (Hou et al., 2021) to detect it, suggests the potential role of (+)-isopinocamphone in the selection of host trees. Kandasamy et al. (2023) tested in a short-range two-choice test in a Petri dish synthetically prepared (+)-isopinocamphone added as a solution in mineral oil to spruce bark agar, which did not exhibit significant attractivity for I. typographus bark beetles in tested doses. This further indicates that the effect of (+)-isopinocamphone we see in trap catches may be a long-range attraction (in accordance with the known long-range attraction of cis-verbenol; Schlyter and Birgersson, 1999).
4.2 Response of bark beetle insect natural enemies to tested compounds
The anticipated captures of the common predatory beetle Thanasimus sp. were relatively low. This could be attributed to the fact that our pheromone bait only contained the two major pheromone components, 2-methyl-3-buten-2-ol and (S)-cis-verbenol, and not ipsdienol, an I. typographus pheromone component emitted in smaller amounts in the later attack states (Birgersson et al., 1984; Hulcr et al., 2006). Furthermore, we observed only a few captures of Medetera sp. and Lonchaea sp. flies, likely due to the use of a trap optimized for Coleoptera that lacked sticky surfaces.
Although there is limited information on Staphylinidae predators using host tree volatiles for locating bark beetles (Wegensteiner et al., 2015), it has been reported that they are attracted to pheromone traps used for monitoring Ips typographus (El-Sayed, 2023). Additionally, commercial pheromone traps tested in combination with host tree logs (P. abies) caught ~38% more predatory Staphylinidae than traps without logs, in comparison to a 32% increase in catches of Thanasimus formicarius (L.) (Zumr, 1983). Hence, host tree compounds may mediate staphylinid prey location.
The pteromalid parasitoid wasp Rhopalicus sp. was the most abundant among the captured bark beetle natural enemies, but there were no significant preferences for any of the tested compounds due to the low number of caught specimens. In the literature, an electroantennographic study of bark beetle gallery smell was tested on the antennae of Rhopalicus tutela (Walker) females. The antennae showed sensitivity to oxygenated monoterpenes and estragole (Pettersson, 2001). Additionally, the olfactory response to estragole was reported in other species of parasitoid wasps, Spathius pallidus Ashmead, 1893, and Roptrocerus xylophagorum (Hymenoptera: Pteromalidae; Sullivan et al., 1997).
5 Conclusion
The effect of 1,8-cineole, estragole, and (+)-isopinocamphone, as observed in our field experiments, provides evidence that these oxygenated monoterpenes and estragole can exhibit biological activity for I. typographus and their natural enemies when combined with I. typographus aggregation pheromone. This suggests that their long-range activity is not solely dependent on a complex mixture, such as that emitted by symbiotic fungi inoculated on wood substrates (Kandasamy et al., 2023).
The discovery of new attraction inhibitors or adjuvants for attractants can be applied to the development of integrated pest management methods for controlling I. typographus. Anti-attractants, a term broadly used for attraction inhibitors, have already been tested to deter various pest bark beetles, such as Dendroctonus ponderosae, D. rufipennis, D. pseudotsugae, Ips pini, and Dryocoetes confusus, from attraction to their pheromone or to the host tree (Schlyter, 2012). These anti-attractants can originate from host trees, non-host trees, associated microorganisms, or the beetles themselves (Borden et al., 2000; Munro et al., 2020).
In the protection of Norway spruce trees against I. typographus, verbenone, a well-established repellent for bark beetles, was tested with varying success (Jakuš et al., 2003; Frühbrodt et al., 2023). In nature, verbenone signals an old and over-exploited host. The synergistic blend effect of verbenone mixed with green leaf volatiles (C6 alcohols) and C8 alcohols (3-octanol and 1-octen-3-ol) and the angiosperm and fungal spiroacetal conophthorin (Zhang and Schlyter, 2004) was also evaluated for tree protection against I. typographus (Schiebe et al., 2011), resulting in a reduction of tree killing ranging from 35 to 76% in protected areas.
The recently tested anti-attractant mixture also includes, besides the 3-octanol, 1-octen-3-ol, hexanol and conophthorin, 1,8-cineole and trans-4-thujanol from spruce and excludes verbenone (Jirošová et al., 2022a). Anti-attractant blends offer partial protection for standing trees but are ineffective for windfallen trees. Adding new anti-attractants, e.g., geranyl acetone (Lindmark et al., 2023), to the mixture may enhance tree protection effects.
A comprehensive approach to semiochemical tree protection against I. typographus attacks could employ the push-pull strategy. Trees are protected by anti-attractants, and repelled beetles are caught in pheromone traps baited with attractive I. typographus pheromones. Both the push and pull might be enhanced by the addition of new semiochemicals (Jakuš et al., 2022; Deganutti et al., 2023). The addition of (+)-isopinocamphone to the trapping lure could increase beetle attraction away from trees while simultaneously protecting them from an estragole-enhanced beetle repellant.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because Ethical review and approval were not required for the study on animals in accordance with the local legislation and institutional requirements. We have performed all beetle experiments that comply with the ARRIVE guidelines and were carried out in accordance with (Scientific Procedures) Act, 1986 and associated guidelines, EU Directive 2010/63/EU for animal experiments.
Author contributions
AACM: Data curation, Formal analysis, Investigation, Validation, Writing—original draft. RJ: Data curation, Investigation, Writing—review & editing. RM: Formal analysis, Writing—review & editing. CRU: Methodology, Writing—review & editing. FS: Conceptualization, Methodology, Supervision, Validation, Writing—review & editing. AJ: Conceptualization, Formal analysis, Investigation, Methodology, Supervision, Validation, Visualization, Writing—original draft, Writing—review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Ministry of Education, Youth and Sport, Operational Programme Research, Development and Education, Czech Republic; “EXTEMIT-K,” No. CZ.02.1.01/0.0/0.0/15_003/0000433. Research funding Internal Grant Commission at the Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Czech Republic; [AACM, IGA: 43950_1312_ 21]. Publication fee funding National Agency for Agricultural Research; Czech Republic NAZV QK1910480.
Acknowledgments
We thank the Forest CZU at Kostelec and Cernými Lesy for permission to conduct experiments on their property and for their support. The authors would like to thank Jaroslav Kašpar from the Military Forests of the Czech Republic and local foresters from the community forest of Liptovský Trnovec Souleymane Diallo and Giorgi Kozhoridze, and Vivek Vikram Sing from the Faculty of Forest Wood Sciences (CZU) and Ondrej Kalina for valuable field assistance. A special thanks to Andrzej Mazur from the Department of Forest Entomology and Pathology of the Faculty of Forestry and Wood Technology, Poznan University of Life Sciences, for assistance with the identification of the Staphylinidae group. Linnaeus University is acknowledged for its support of CRU.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2023.1292581/full#supplementary-material
References
Andersson, M. N. (2012). Mechanisms of odor coding in coniferous bark beetles: from neuron to behavior and application. Psyche (Camb. Mass.). 2012, 1-14. doi: 10.1155/2012/149572
Andersson, M. N., Larsson, M. C., BlaŽenec, M., Jakuš, R., Zhang, Q. H., and Schlyter, F. (2010). Peripheral modulation of pheromone response by inhibitory host compound in a beetle. J. Exp. Biol. 19, 3332-3339. doi: 10.1242/jeb.044396
Andersson, M. N., Larsson, M. C., and Schlyter, F. (2009). Specificity and redundancy in the olfactory system of the bark beetle Ips typographus: single-cell responses to ecologically relevant odors. J. Insect Physiol. 55, 556-567. doi: 10.1016/j.jinsphys.2009.01.018
Arnett, Jr, R. H., and Thomas, M. C. (2000). American Beetles: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. Boca Raton, FL: CRC Press.
Bakke, A., and Kvamme, T. (1981). Kairomone response in Thanasimus predators to pheromone components of Ips typographus. J. Chem. Ecol. 7, 305–312. doi: 10.1007/BF00995753
Bergström, G. (2007). Chemical ecology= chemistry+ ecology! Pure Appl. Chem. 12, 2305-2323. doi: 10.1351/pac200779122305
Binyameen, M., Jankuvová, J., BlaŽenec, M., Jakuš, R., Song, L., Schlyter, F., et al. (2014). Co-localization of insect olfactory sensory cells improves the discrimination of closely separated odour sources. Funct. Ecol. 5, 1216-1223. doi: 10.1111/1365-2435.12252
Birgersson, G., Schlyter, F., Löfqvist, J., and Bergström, G. (1984). Quantitative variation of pheromone components in the spruce bark beetle Ips typographus from different attack phases. J. Chem. Ecol. 10, 1029-1055. doi: 10.1007/BF00987511
BlaŽyte-Cereškiene, L., Apšegaite, V., RadŽiute, S., Mozuraitis, R., Buda, V., and Pečiulyte, D. (2016). Electrophysiological and behavioural responses of Ips typographus (L.) to trans-4-thujanol—a host tree volatile compound. Ann. For. Sci. 2, 247-256. doi: 10.1007/s13595-015-0494-5
Blomquist, G. J., Tittiger, C., MacLean, M., and Keeling, C. I. (2021). Cytochromes P450: terpene detoxification and pheromone production in bark beetles. Curr. Opin. Insect Sci. 43, 97-102. doi: 10.1016/j.cois.2020.11.010
Borden, J. H., Chong, L. J., Gries, G., Gries, R., Huber, D. P., Pierce, Jr. H. D., et al. (2000). Non-Host Volatiles as Repellents for Conifer-Infesting Bark Beetles. U.S. Patent 6,051,612. Washington, DC: U.S. Patent and Trademark Office.
Bouček, Z., and Rasplus, J. Y. (1991). Illustrated key to West-Palearctic genera of Pteromalidae (Hymenoptera: Chalcidoidea). Paris: Editions Quae Press; Institut National de la Recherche Agronomique (INRA).
Brázdil, R., Zahradník, P., Szabó, P., Chromá, K., Dobrovolný, P., Dolák, L., et al. (2022). Meteorological and climatological triggers of notable past and present bark beetle outbreaks in the Czech Republic. Clim. Past 9, 2155-2180. doi: 10.5194/cp-18-2155-2022
Brooks, M. E., Kristensen, K., Van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2, 378-400. doi: 10.32614/RJ-2017-066
Celedon, J. M., and Bohlmann, J. (2019). Oleoresin defenses in conifers: chemical diversity, terpene synthases and limitations of oleoresin defense under climate change. New Phytol. 224, 1444–1463. doi: 10.1111/nph.15984
Chiu, C. C. (2018). Detoxification of Pine Terpenoids by Mountain Pine Beetle Cytochromes P450 (T). University of British Columbia. Available online at: https://open.library.ubc.ca/collections/ubctheses/24/items/1.0376017 (accessed April 28, 2023).
Chiu, C. C., Keeling, C. I., and Bohlmann, J. (2018). Monoterpenyl esters in juvenile mountain pine beetle and sex-specific release of the aggregation pheromone trans-verbenol. Proc. Natl. Acad. Sci. U. S. A. 14, 3652-3657. doi: 10.1073/pnas.1722380115
Chiu, C. C., Keeling, C. I., and Bohlmann, J. (2019). The cytochrome P450 CYP6DE1 catalyzes the conversion of α-pinene into the mountain pine beetle aggregation pheromone trans-verbenol. Sci. Rep. 9, 1-1477. doi: 10.1038/s41598-018-38047-8
Dai, L., Gao, H., and Chen, H. (2021). Expression levels of detoxification enzyme genes from Dendroctonus armandi (Coleoptera: Curculionidae) fed on a solid diet containing pine phloem and terpenoids. Insects 12, 926. doi: 10.3390/insects12100926
Deganutti, L., Biscontin, F., Bernardinelli, I., and Faccoli, M. (2023). The semiochemical push-and-pull technique can reduce bark beetle damage in disturbed Norway spruce forests affected by the Vaia storm. Agric. For. Entomol. 2023, 12600. doi: 10.1111/afe.12600
El-Sayed, A. M. (2023). The Pherobase: Database of Pheromones and Semiochemicals. Available online at: http://www.pherobase.com/database/family/family-Staphylinidae.php (accessed June 12, 2023).
Erbilgin, N., and Raffa, K. F. (2001). Modulation of predator attraction to pheromones of two prey species by stereochemistry of plant volatiles. Oecologia 127, 444-453. doi: 10.1007/s004420000606
Evans, A. B., Martin, N. G., Minden, K., and Ollis, M. A. (2020). Infinite latin squares: neighbor balance and orthogonality. Electron. J. Comb. 27, 53. doi: 10.37236/8020
Fettig, C. J., and Hilszczański, J. (2015). “Management strategies for bark beetles in conifer forests,” in Bark Beetles: Biology and Ecology of Native and Invasive Species, eds E. F. Vega and W. R. Hofstetter (Amsterdam: Elsevier Inc.), 555-584.
Frühbrodt, T., Schebeck, M., Andersson, M. N., Holighaus, G., Kreuzwieser, J., Burzlaff, T., et al. (2023). Verbenone—the universal bark beetle repellent? Its origin, effects, and ecological roles. J. Pest Sci. 31, 1-37. doi: 10.1007/s10340-023-01635-3
Ganji, S., Svensson, F. G., and Unelius, C. R. (2020). Asymmetric synthesis of oxygenated monoterpenoids of importance for bark beetle ecology. J. Nat. Prod. 83, 3332-3337. doi: 10.1021/acs.jnatprod.0c00669
Ghimire, R. P., Kivimäenpää, M, Blomqvist, M., Holopainen, T., Lyytikäinen-Saarenmaa, P., and Holopainen, J. K. (2016). Effect of bark beetle (Ips typographus L.) attack on bark VOC emissions of Norway spruce (Picea abies Karst.) trees. Atmos. Environ. 1, 145-152. doi: 10.1016/j.atmosenv.2015.11.049
Graham, M. W. R. V. (1969). The Pteromalidae of North-Western Europe (Hymenoptera: Chalcidoidea). Bulletin of the British Museum (Natural History). London: London Press.
Haack, R. A., Lawrence, R. K., Petrice, T. R., and Poland, T. M. (2004). Disruptant effects of 4-allylanisole and verbenone on Tomicus piniperda (Coleoptera: Scolytidae) response to baited traps and logs. Gt. Lakes Entomol. 37, 4. doi: 10.22543/0090-0222.2107
Hallberg, E. (1982). Sensory Organs in Ips typographus (Insecta: Coleoptera) Fine structure of the sensilla of the maxillary and labial palps. Acta Zool. 63, 191-198. doi: 10.1111/j.1463-6395.1982.tb00778.x
Hardin, J. W., and Hilbe, J. M. (2007). Generalized Linear Models and Extensions. College Station, TX: Stata press.
Hayes, J. L., and Strom, B. L. (1994). 4-Allylanisole as an inhibitor of bark beetle (Coleoptera: Scolytidae) aggregation. J. Econ. Entomol. 87, 1586-1594. doi: 10.1093/jee/87.6.1586
Heber, T., Helbig, C. E., Osmers, S., and Müller, M. G. (2021). Evaluation of attractant composition, application rate, and trap type for potential mass trapping of Ips typographus (l.). Forests. 12, 1727. doi: 10.3390/f12121727
Hlásny, T., Barka, I., Merganičová, K., Krístek, Š., Modlinger, R., Turčáni, M., et al. (2022). A new framework for prognosing forest resources under intensified disturbance impacts: case of the Czech Republic. For. Ecol. Manag. 523, 120483. doi: 10.1016/j.foreco.2022.120483
Hlásny, T., Krokene, P., Liebhold, A., and Montagné-Huck, C. (2019). Living With Bark Beetles: Impacts, Outlook and Management Options. Finland: European Forest Institute.
Hou, X. Q., Yuvaraj, J. K., Roberts, R. E., Zhang, D. D., Unelius, C. R., Löfstedt, C., et al. (2021). Functional evolution of a bark beetle odorant receptor clade detecting monoterpenoids of different ecological origins. Mol. Biol. Evol. 38, 4934-4947. doi: 10.1093/molbev/msab218
Hulcr, J., Ubik, K., and Vrkoc, J. (2006). The role of semiochemicals in tritrophic interactions between the spruce bark beetle Ips typographus, its predators and infested spruce. J. Appl. Entomol. 130, 275-283. doi: 10.1111/j.1439-0418.2006.01069.x
Jaakkola, E., Gärtner, A., Jönsson, A. M., Olsson, K. L. P., and Holst, T. (2022). Spruce bark beetle (Ips typographus) infestation cause up to 700 times higher bark BVOC emission rates from Norway spruce (Picea abies). Biogeosciences 20, 803–826. doi: 10.5194/bg-2022-125
Jakuš, R., Modlinger, R., Kašpar, J., Majdak, A., BlaŽenec, M., Korolyova, N., et al. (2022). Testing the efficiency of the push-and-pull strategy during severe Ips typographus outbreak and extreme drought in Norway Spruce Stands. Forests 13, 2175. doi: 10.3390/f13122175
Jakuš, R., Schlyter, F., Zhang, Q. H., BlaŽenec, M., Vaverčák, R., Grodzki, W., et al. (2003). Overview of development of an anti-attractant based technology for spruce protection against Ips typographus: from past failures to future success. Anzeiger für Schädlingskunde. J. Pest Sci. 76, 89-99. doi: 10.1046/j.1439-0280.2003.03020.x
Jirošová, A., Jakuš, R, Modlinger, R., Turcani., M, and Schlyter, F. (2022a). “Prípravek pro repelenci lýkoŽrouta smrkového“ 309426 Úrad prumyslového vlastnictví, Patent CR.
Jirošová, A., Kalinová, B., Modlinger, R., Jakuš, R., Unelius, C. R., BlaŽenec, M., et al. (2022b). Anti-attractant activity of (+)-trans-4-thujanol for Eurasian spruce bark beetle Ips typographus: novel potency for females. Pest Manag. Sci. 78, 1992-1999. doi: 10.1002/ps.6819
Joe, H., and Zhu, R. (2005). Generalized poisson distribution: the property of mixture of Poisson and comparison with negative binomial distribution. J. Math. Biosci. 47, 219–229. doi: 10.1002/bimj.200410102
Joseph, G., Kelsey, R. G., Peck, R. W., and Niwa, C. G. (2001). Response of some scolytids and their predators to ethanol and 4-allylanisole in pine forests of central Oregon. J. Chem. Ecol. 27, 697-715. doi: 10.1023/A:1010345817756
Kalinová, B., Brízová, R., KníŽek, M., Turčáni, M., and Hoskovec, M. (2014). Volatiles from spruce trap-trees detected by Ips typographus bark beetles: chemical and electrophysiological analyses. Arthropod-Plant Interact. 8, 305-316. doi: 10.1007/s11829-014-9310-7
Kandasamy, D., Zaman, R., Nakamura, Y., Zhao, T., Hartmann, H., Andersson, M. N., et al. (2021). Bark beetles locate fungal symbionts by detecting volatile fungal metabolites of host tree resin monoterpenes. BioRxiv 4, 2021-2007. doi: 10.1101/2021.07.03.450988
Kandasamy, D., Zaman, R., Nakamura, Y., Zhao, T., Hartmann, H., Andersson, M. N., et al. (2023). Conifer-killing bark beetles locate fungal symbionts by detecting volatile fungal metabolites of host tree resin monoterpenes. PLoS Biol. 21, 3001887. doi: 10.1371/journal.pbio.3001887
Krokene, P. (2015). “Conifer defense and resistance to bark beetles,” in Bark Beetles, eds F. E. Vega and R. W. Hofstetter (San Diego, CA: Academic Press), 177–207. doi: 10.1016/B978-0-12-417156-5.00005-8
Lehmanski, L. M., Kandasamy, D., Andersson, M. N., Netherer, S., Alves, E. G., Huang, J., et al. (2023). Addressing a century old hypothesis–do pioneer beetles of Ips typographus use volatile cues to find suitable host trees?. New Phytol. 238, 1762–1770. doi: 10.1111/nph.18865
Lindmark, M., Ganji, S., Wallin, E. A., Schlyter, F., and Unelius, C. R. (2023). Semiochemicals produced by fungal bark beetle symbiont Endoconidiophora rufipennis and the discovery of an anti-attractant for Ips typographus. PLoS ONE 18, 4. doi: 10.1371/journal.pone.0283906
Munro, H. L., Gandhi, K. J., Barnes, B. F., Montes, C. R., Nowak, J. T., Shepherd, W. P., et al. (2020). Electrophysiological and behavioral responses Dendroctonus frontalis and D. terebrans (Coleoptera: Curculionidae) to resin odors of host pines (Pinus spp.). Chemoecology 30, 215-231. doi: 10.1007/s00049-020-00311-7
Navarrete, J. L., and Newton, A. F. (2003). Guía ilustrada para los géneros de Staphylinidae (Coleoptera) de México. Biota Colomb. 4, 271–273.
Netherer, S., Kandasamy, D., Jirosová, A., Kalinová, B., Schebeck, M., and Schlyter, F. (2021). Interactions among Norway spruce, the bark beetle Ips typographus and its fungal symbionts in times of drought. J. Pest Sci. 94, 591–614. doi: 10.1007/s10340-021-01341-y
Peck, O., Bouček, Z., and Hoffer, A. (1964). Keys to the Chalcidoidea of Czechoslovakia (Insecta: Hymenoptera). Mem. Ent. Soc. Can. 96, 7–121.
Pelto-Arvo, M. (2020). The Impact of Forest Health Status on Natural Enemies and Associates of the European Spruce Bark Beetle Ips typographus (L.). (Master‘s thesis), University of Helsinki, Helsinki, Finland.
Pettersson, E. M. (2001). Volatiles from potential hosts of Rhopalicus tutela a bark beetle parasitoid. J. Chem. Ecol. 27, 2219-2231. doi: 10.1023/A:1012226820035
Pettersson, E. M., and Boland, W. (2003). Potential parasitoid attractants, volatile composition throughout a bark beetle attack. Chemoecology 13, 27-37. doi: 10.1007/s000490300003
R Core Team (2023). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Raffa, K. F. (2014). Terpenes tell different tales at different scales: glimpses into the chemical ecology of conifer-bark beetle-microbial interactions. J. Chem. Ecol. 40:1–20. doi: 10.1007/s10886-013-0368-y
Ramakrishnan, R., Hradecký, J., Roy, A., Kalinová, B., Mendezes, R. C., Synek, J., et al. (2022). Metabolomics and transcriptomics of pheromone biosynthesis in an aggressive forest pest Ips typographus. Insect Biochem. Mol. Biol. 140, 103680. doi: 10.1016/j.ibmb.2021.103680
Schebeck, M., Schopf, A., Ragland, G. J., Stauffer, C., and Biedermann, P. H. (2023). Evolutionary ecology of the bark beetles Ips typographus and Pityogenes chalcographus. Bull. Entom. Res. Lond. 113, 1-0. doi: 10.1017/S0007485321000353
Schiebe, C., BlaŽenec, M., Jakuš, R., Unelius, C. R., and Schlyter, F. (2011). Semiochemical diversity diverts bark beetle attacks from Norway spruce edges. J. Appl. Entomol. 135, 726-737. doi: 10.1111/j.1439-0418.2011.01624.x
Schiebe, C., Hammerbacher, A., Birgersson, G., Witzell, J., Brodelius, P. E., Gershenzon, J., et al. (2012). Inducibility of chemical defenses in Norway spruce bark is correlated with unsuccessful mass attacks by the spruce bark beetle. Oecologia 170, 183-198. doi: 10.1007/s00442-012-2298-8
Schiebe, C., Unelius, C. R., Ganji, S., Binyameen, M., Birgersson, G., and Schlyter, F. (2019). Styrene, (+)-trans-(1R,4S,5S)-4-Thujanol and oxygenated monoterpenes related to host stress elicit strong electrophysiological responses in the bark beetle Ips typographus. J. Chem. Ecol. 45, 474–489. doi: 10.1007/s10886-019-01070-8
Schlyter, F. (2012). Semiochemical diversity in practice: antiattractant semiochemicals reduce bark beetle attacks on standing trees—a first meta-analysis. Psyche (Camb. Mass.) 2012, 268621. doi: 10.1155/2012/268621
Schlyter, F., and Birgersson, G. (1999). “Forest beetles,” in Pheromones of Non-Lepidopteran Insects Associated With Agricultural Plants, eds J. Hardie and A. K. Minks (Wallingford, CT: CABI Publishing), 113-148.
Sousa, M., Birgersson, G., Karlsson Green, K., Pollet, M., and Becher, P. G. (2023). Odors attracting the long-legged predator Medetera signaticornis Loew to Ips typographus L. Infested Norway Spruce Trees. J. Chem. Ecol. 6, 1-14. doi: 10.1007/s10886-023-01405-6
Sullivan, B. T., Berisford, C. W., and Dalusky, M. J. (1997). Field response of southern pine beetle parasitoids to some natural attractants. J. Chem. E. 23, 837-856. doi: 10.1023/B:JOEC.0000006415.17532.d0
Wegensteiner, R., Wermelinger, B., and Herrmann, M. (2015). “Natural enemies of bark beetles: predators, parasitoids, pathogens, and nematodes,” in Bark Beetles: Biology and Ecology of Native and Invasive Species, eds E. F. Vega and W. R. Hofstetter (Amsterdam: Elsevier Inc.), 246-304.
Werner, R. A. (1995). Toxicity and repellency of 4–allylanisole and monoterpenes from white spruce and tamarack to the spruce beetle and eastern larch beetle (Coleoptera: Scolytidae). Environ. Entomol. 1, 24372-24379. doi: 10.1093/ee/24.2.372
Zhang, Q. H., and Schlyter, F. (2004). Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric. For. Entomol. 202, 1-20. doi: 10.1111/j.1461-9555.2004.00202.x
Zumr, V. (1983). Effect of synthetic pheromones Pheroprax on the coleopterous predators of the spruce bark beetle Ips typographus (L.). Zeit. A. Entomol. 95, 47-50. doi: 10.1111/j.1439-0418.1983.tb02609.x
Keywords: Eurasian spruce bark beetle, host compounds, Pteromalidae, Staphylinidae, Norway spruce, Picea abies, spruce kairomone, pheromone traps
Citation: Moliterno AAC, Jakuš R, Modlinger R, Unelius CR, Schlyter F and Jirošová A (2023) Field effects of oxygenated monoterpenes and estragole combined with pheromone on attraction of Ips typographus and its natural enemies. Front. For. Glob. Change 6:1292581. doi: 10.3389/ffgc.2023.1292581
Received: 11 September 2023; Accepted: 22 November 2023;
Published: 21 December 2023.
Edited by:
Davide Rassati, University of Padua, ItalyReviewed by:
Nadir Erbilgin, University of Alberta, CanadaBrian Sullivan, Southern Research Station, Forest Service (USDA), United States
Copyright © 2023 Moliterno, Jakuš, Modlinger, Unelius, Schlyter and Jirošová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anna Jirošová, jirosovaa@fld.czu.cz
†These authors have contributed equally to this work