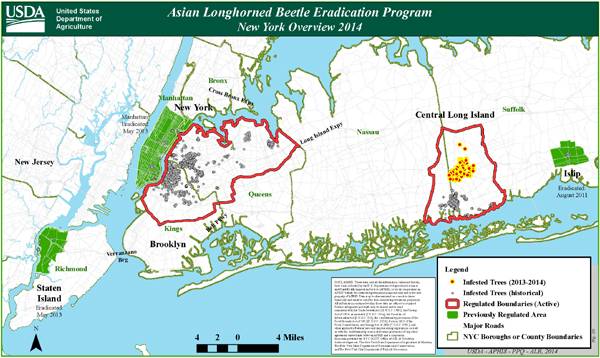
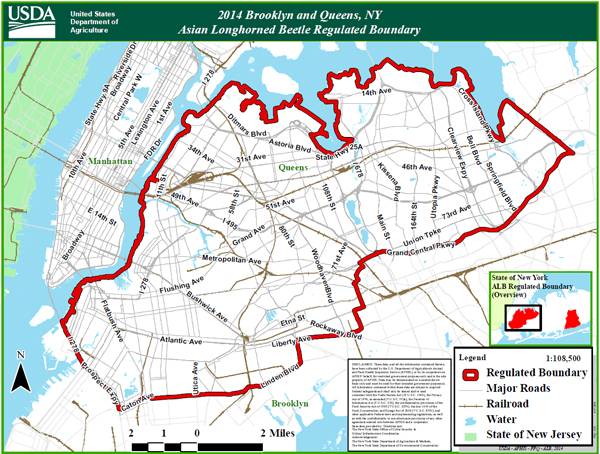
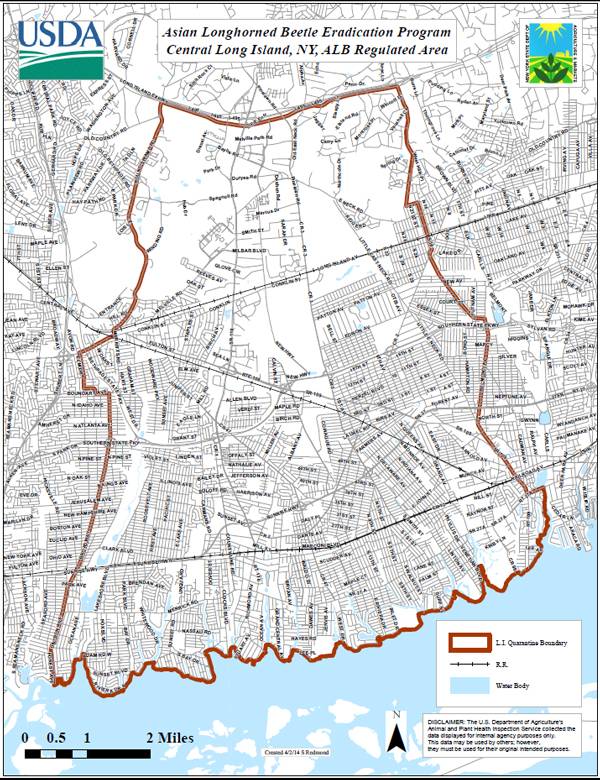
Problem | History | Biology | Habitat | Management | New York Distribution Map

Problem
Mile-a-minute weed (Persicaria perfoliata) is a vigorous, barbed vine that smothers other herbaceous plants, shrubs and even trees by growing over them. Growing up to six inches per day, mile-a-minute weed forms dense mats that cover other plants and then stresses and weakens them through smothering and physically damaging them. Sunlight is blocked, thus decreasing the covered plant’s ability to photosynthesize; and the weight and pressure of the mile-a-minute weed can cause poor growth of branches and foliage. The smothering can eventually kill overtopped plants.
History
Mile-a-minute weed (Persicaria perfoliata (L.) H. Gross, formerly Polygonum perfoliatum) is a member of the polygonum or buckwheat family. It is native to India and Eastern Asia and was accidentally introduced via contaminated holly seed into York County, Pennsylvania in 1930. Mile-a-minute weed has been found in all the Mid-Atlantic states, southern New England, North Carolina, Ohio, and Oregon (2011). In New York, mile-a-minute weed has been recorded mostly in counties south of the northern Connecticut border. Mile-a-minute weed has a large potential to expand in cooler areas, as the seed requires an eight-week cold period in order to flower. It is estimated that mile-a-minute weed is in only 20% of its potential U.S. range.
Infestations of mile-a-minute weed decrease native vegetation and habitat in natural areas impacting plants and the wildlife that depend on those plants as well. Mile-a-minute weed can also be a major pest in Christmas tree plantations, reforestation areas and young forest stands, and landscape nurseries. Areas that are regularly disturbed, such as powerline and utility right-of-ways where openings are created through regular herbicide use are prime locations for mile-a-minute weed establishment. Small populations of rare plants could be completely destroyed. Thickets of these barbed plants can also be a deterrent to recreation.
Biology
Mile-a-minute weed is an herbaceous annual vine. Its leaves are alternate, light green, 4 to 7 cm long and 5 to 9 cm wide, and shaped like an equilateral triangle. Its green vines are narrow and delicate, becoming woody and reddish with time. The vines and the undersides of leaves are covered with recurved barbs that aid in its ability to climb. Mile-a-minute has ocreae that surround the stems at nodes. This distinctive 1 to 2 cm feature is cup-shaped and leafy. Flower buds, and thus flowers and fruit, grow from these ocreae. When the small, white, inconspicuous flowers are pollinated they form spikes of blue, berry-like fruits, each containing a single glossy, black seed called an achene. Vines can grow up to six inches per day.

Mile-a-minute weed is primarily a self-fertile plant and does not need any pollinators to produce viable seeds. Its ability to flower and produce seeds over a long period of time (June through October) make mile-a-minute weed a prolific seeder. Seeds can be viable in the soil for up to six years and can germinate at staggered intervals. Vines are killed by frost and the seeds overwinter in the soil. Mile-a-minute seeds require an eight-week vernalization period at temperatures below 10 degrees Celsius in order to flower, and therefore be a threat. Germination is generally early April through early July.
Seeds are carried long distances by birds, which are presumed to be the main cause of long distance spread. Deer, chipmunks, squirrels and even one particular species of ant is known to eat mile-a-minute weed fruit. Viable seeds have been found in deer scat; an indication that other animals may also be vectors.
Mile-a-minute weed seeds can float for seven to nine days, which allows for long distance movement in water. This movement can be amplified during storms when vines hanging over waterways drop their fruit into fast moving waters, which then spread the seeds throughout a watershed.
Habitat
Mile-a-minute weed is generally found colonizing natural and man-made disturbed and open areas and along the edges of woods, streams, wetlands, uncultivated fields, and roads. It can also be found in areas with extremely wet environments with poor soil structure, and while it will grow in drier soils, mile-a-minute prefers high moisture soils. It will tolerate some shade for part of the day, but prefers full sun. Using its specially-adapted recurved barbs, mile-a-minute weed can reach sunlight by climbing over plants, helping it outcompete other vegetation.

Management
Mile-a-minute has a number of management options that can be employed. Different sites will dictate different levels of management depending on conditions and the level of infestation. Once all the plants have been removed, on-going monitoring and management must occur for up to six years in order to exhaust any seeds remaining in the soil.
Biological Control
The mile-a-minute weevil, Rhinocominus latipes Korotyaev, is a 2 mm long, black weevil which is often covered by an exuded orange film produced from the mile-a-minute plants it feeds on. This small weevil is host-specific to mile-a-minute weed and has been successfully released and recovered in multiple locations in the U.S.

The adult weevils feed on the leaves of mile-a-minute weed and females lay eggs on the leaves and stems. When the eggs hatch, the larvae bore into the stem to complete their development, feeding on the stems between the nodes. The larvae then emerge and drop to the soil to pupate. There are three to four overlapping generations per year, with about a month needed per generation. Egg laying ceases in late summer or early fall, and the mile-a-minute weevil overwinters as an adult in the soil or leaf litter.
Mile-a-minute weevil feeding damage can stunt plants by causing the loss of apical dominance and can delay seed production. In the presence of competing vegetation, mile-a-minute weed can be killed by the weevil. The mile-a-minute weevil is more effective in the sun than in the shade. Over time, mile-a-minute weevils have been shown to reduce spring seedling counts. Biological control of mile-a-minute weed is currently the most promising and cost effective method.

For more information on the mile-a-minute weevil, check the University of Delaware Biological Control on Invasive Plants Research website:
Cultural Control
Cultural methods can be used to help prevent mile-a-minute weed introduction to a new area. Maintain a stable plant community; avoid creating disturbances, openings or gaps in existing vegetation; and maintain wide, shade-producing, vegetative buffers along streams and wooded areas to prevent establishment.
Manual and Mechanical Control
Hand-pulling of vines can be effective; ideally before the barbs harden, afterwards thicker gloves are needed. Pull and bale vines and roots as early in the season as possible. Let the piles of vines dry out completely before disposing. Later in the season, vines must be pulled with caution as the fruit could be knocked off or spread more easily. Collected plants can be incinerated or burned, left to dry and piled on site, or bagged and landfilled (least preferred). Dry piles left on site should be monitored and managed a few times each year, especially during the spring and early summer germination period to ensure any germinating seedlings are destroyed.
Low growing populations of mile-a-minute weed can have their resources exhausted through repeated mowing or cutting. This will reduce flower production and therefore reduce fruit production.
Chemical Control
Mile-a-minute weed can be controlled with commonly used herbicides in moderate doses. The challenge with herbicides is mile-a-minute’s ability to grow over the top of desirable vegetation, and spraying the foliage of only the mile-a-minute weed can be challenging. Pre-emergent herbicides (herbicides that prevent seed germination) can be used with extensive infestations, often in combination with spot treatments of post-emergent herbicides (herbicides applied to the growing plant) for seedlings that escape control. Small populations are better controlled with post-emergent herbicides. General chemical control guidelines can be found at http://www.docs.dcnr.pa.gov/cs/groups/public/documents/document/dcnr_20033415.pdf. Areas treated with herbicides need to be monitored and retreated as necessary when new seedlings emerge from the seed bank, see above. Please contact your local Cornell Cooperative Extension office http://www.cce.cornell.edu for pesticide use guidelines. For treating wetland areas or infestations near water, contact a certified pesticide applicator. Always apply pesticides according to the label directions; it’s the law.
New York Distribution Map
This map shows confirmed observations (green points) submitted to the NYS Invasive Species Database. Absence of data does not necessarily mean absence of the species at that site, but that it has not been reported there. For more information, please visit iMapInvasives.































.jpg)
.jpg)